 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 段勇杰, 谢婷, 高云涛, 王枫林, 李梦迪, 关长涛, 贾玉东. 2023.
- DUAN Yong-Jie, XIE Ting, GAO Yun-Tao, WANG Feng-Lin, LI Meng-Di, GUAN Chang-Tao, JIA Yu-Dong. 2023.
- 斑石鲷(Oplegnathus punctatus)游泳性能研究
- STUDY ON SWIMMING ABILITY OF SPOTTED KNIFEJAW (OPLEGNATHUS PUNCTATUS)
- 海洋与湖沼, 54(1): 183-193
- Oceanologia et Limnologia Sinica, 54(1): 183-193.
- http://dx.doi.org/10.11693/hyhz20220400106
文章历史
-
收稿日期:2022-04-24
收修改稿日期:2022-07-09




2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 山东青岛 266237;
3. 中国水产科学研究院黄海水产研究所 山东青岛 266071;
4. 鲁东大学农学院 山东烟台 264025
2. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology(Qingdao), Qingdao 266237, China;
3. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China;
4. College of Agriculture, Ludong University, Yantai 264025, China
目前, 随着我国近海养殖环境逐渐恶化, 野生渔业资源日益枯竭, 发展深远海养殖对于解决诸多实际问题具有重要意义, 深远海养殖发展不仅要考虑养殖基础设施建设, 还要建立养殖鱼类游泳能力评价体系, 筛选深远海海区适合养殖的品种规格(徐琰斐等, 2021; 徐皓等, 2021)。游泳能力是水生生物生存与适应进化的重要基础, 大多数鱼类通过游泳活动进行捕食、求偶、迁徙和逃避敌害等(Kaufmann et al, 1992)。Yanase等(2007)提出了游泳时间和游泳速度是评价鱼类游泳能力的重要指标, 并对其进行了进一步分类与探讨, 分为续航游泳时间、临界游泳速度和暴发游泳速度等(Watkins, 1996; Plaut, 2000; Yanase et al, 2007)。在游泳过程中, 鱼类会消耗氧气, 产生能量消耗, 影响其游泳运动能量的分配和选择, 以运动耗氧率表示(Arendt et al, 2005; Farrell, 2008), 鱼类的游泳效率是指单位体重的鱼类移动单位距离所消耗的能量, 以单位位移耗能表示(Ylieff et al, 2003; Fu et al, 2009), 单位位移耗能曲线随着游泳速度的增加呈逐渐下降至平稳趋势, 曲线存在一个最低点, 处于该点时鱼体的单位位移耗能最少, 能量效率最高, 其所对应的游泳速度被称为最适游泳速度(Peterson et al, 1969; Westerterp, 1977; Mehner et al, 1994)。目前, 有关鱼类游泳能力研究主要集中在淡水鱼类, 研究多以设计鱼道设施、保护鱼类栖息地和渔业资源为目的, 同时为鱼类行为生态学的相关理论研究提供参考资料(倪海儿等, 2009)。以斑重唇鱼(Diptychus maculatus Steindachner)为鱼道主要过鱼对象时, 为促进鱼体顺利进入鱼道, 建议设计的进口流速为1.02~1.39 m/s, 休息池主流为0.20~1.02 m/s, 鱼道竖缝处流速宜低于0.85 m/s, 雷青松等(2020)以暗色唇鲮(Semilabeo obscurus)、华南鲤(Cyprinus carpio rubrofuscus)和红魾(Bagarius rutilus)为鱼道主要过鱼对象时, 建议过鱼设施的内部流速不应小于0.1 m/s, 入口的设计流速范围为0.6~1.0 m/s, 休息区域的流速范围为0.1~0.6 m/s, 孔口或竖缝等高流速区的设计流速不宜超过0.8 m/s (王永猛等, 2021), Richer等(2020)通过对九种鱼类进行游泳能力测定实验, 得出评估各种鱼道类型的设计标准(如坡度、容量、粗糙度和配置)和性能, Hoover等(2017)取得成年大头鱼和鲢鱼的游泳性能数据, 通过对比来评估和优化鱼道结构。虽然海水养殖鱼类的游泳性能研究较少, 但鱼类的耐流特性和养殖业的关系已开始引起人们的关注(王萍等, 2010a), 王萍等(2010a)对美国红鱼(Sciaenops ocellatus)、鲈鱼(Lateolabrax maculatus)、斜带髭鲷(Hapalogenys nitens)进行耐流实验, 筛选出海区适宜的养殖品种。Johansson等(2014)发现网箱中的大西洋鲑(Salmo salar)在不同流速下会产生相应的群体游泳行为, 并得出该种鱼种在网箱养殖中耗能最小的最适巡航速度; Nobata等(2022)通过对太平洋鲑(Oncorhynchus nerka)苗种进行暴发游泳能力测定研究, 发现通过加强游泳运动可以显著改善苗种的质量。鱼类游泳性能即是改进养殖设施设计和建造的重要依据, 也有助于明确深远海养殖品种最适放养规格, 选择适合的养殖海区, 科学放养, 进而促进深远海养殖高质量有序发展(许燕等, 2018)。
斑石鲷(Oplegnathus punctatus), 属鲈形目(Perciformes)、石鲷(Oplegnathidae)、石鲷属(Oplegnathus), 俗称斑鲷、花金鼓、黑金鼓, 主要分布于朝鲜, 日本及中国台湾省、南海、东海、黄海等海域, 在自然海域中, 斑石鲷的自然资源很少, 极少能形成渔汛(孟乾等, 2020; 赵玉柱等, 2021)。作为海水养殖的重要品种, 斑石鲷在深远海养殖过程中具有生长速度快、养殖成本低、抗性强等优势(董登攀等, 2010), 目前关于斑石鲷的研究主要集中在繁殖、生理、病害等方面, 关于游泳性能方面的研究尚未见报道(Wang et al, 2021), Xu等(2022)通过对不同样本转录组测序分析, 初步研究斑石鲷对MCV病毒感染的免疫反应; 赵玉柱等(2021)发现斑石鲷在细胞学分化时间上精巢和卵巢大致同步, 且性腺发育为雌雄异体的分化类型。此外, 斑石鲷在深远海大型工程化围栏养殖模式下, 其生长性状和生理状态相较于工厂化养殖有显著提升(Jia et al, 2021)。流速是影响陆海接力放养规格的关键因素, 明确斑石鲷的耐流能力, 对于制定科学放养时序, 指导陆基工厂化和深远海工程化围栏及网箱养殖, 为陆海接力科学高效放养提供技术支撑。基于此, 本文研究了不同规格斑石鲷的续航游泳时间、暴发游泳速度、临界游泳速度、运动耗氧率, 计算推导出单位位移耗能和最适游泳速度, 并讨论分析无氧运动能力和有氧运动能力之间的关联, 筛选出抗流和非抗流规格, 将为斑石鲷陆海接力衔接规格的选择提供参考依据, 进一步完善陆海接力高效养殖技术工艺。
1 材料与方法 1.1 实验材料实验用斑石鲷(共108尾)购自烟台明波水产有限公司, 根据不同规格和实验需求分别放置循环水养殖水槽中暂养2~3 d, 暂养水体均取自沉淀净化后的天然海水。暂养期间, 通过充氧泵(渔亭ACO_008)保持水槽氧气充足, 水体温度控制在(24.00±1.08) ℃, 每日9:00和16:00饱食定时投喂商业饲料(海童, 潍坊三通生物公司, 饲料中水分、粗蛋白和粗脂肪含量占比分别为8.02%、51.84%、12.50%), 并及时清理残饵和粪便。暂养结束后, 选取健康活泼、体型相近的实验鱼进行实验, 实验用鱼为四种不同规格[体重(50.00±6.00)、(150.00±10.00)、(250.00±14.00)、(350.00±18.00) g, 对应体长分别为(10.00±0.28)、(12.00±0.45)、(16.00±0.84)、(21.00±1.14) cm], 为避免测量体重、体长对实验鱼产生影响, 上述数据均为实验后测量的结果, 每项指标测定实验重复进行四次。
1.2 实验仪器与设备实验设备为本团队自行设计, 委托青岛海星仪器有限公司组装, 并应用于测试鱼类游泳能力相关研究(图 1)。设备主体由内部和外部两个部分构成, 内部为封闭实验区域, 盛水体积为87 L, 外部为水槽, 手动开关交换海水, 材质分别为亚克力和钢化玻璃, 全长145 cm, 鱼类游泳观测视窗尺寸60 cm×20 cm, 传动系统由螺旋桨、电机(SIEMENS/IMB3)和变频器(海利普/1D521)组成。溶解氧测定仪(Aqua TROLL/600)和流速仪(风途/LS20B)测量于探头插孔处, 实验设备上部和鱼类游泳观测视窗用摄像机全程记录。
 |
| 图 1 鱼类游泳能力及运动消耗测定装置 Fig. 1 Measuring device for swimming ability and sports consumption of fish 注: 1. 探头插孔, 2. 整流器, 3. 水流方向, 4. 游泳区, 5. 螺旋桨, 6. 电机, 7. 变频器 |
根据线性拟合Y=0.170 3x+0.029 4, R2=0.998 (见图 2), 实验装置有较高的传动效率, 通过控制变频器频率, 改变螺旋桨转速, 实现装置内水流速度大小变化, 调校变频器频率与流速有较好的线性关系, 造流可控流速范围0~1.95 m/s, 流速由0 m/s增至1.95 m/s的加速时间小于6 s, 装置内有弧形水流通道和整流器, 流速均匀稳定。
 |
| 图 2 流速与变频器频率关系 Fig. 2 Relationship between flow rate and frequency converter |
实验预设方案制定五组流速水平, 分别为50、60、70、80、90、100 cm/s, 在预实验的过程中发现, 四种规格实验鱼可以在50 cm/s流速下进行长时间游泳(> 9 h), 在100 cm/s流速下游泳时间不足10 s, 被水流直接冲击到隔网, 因此最后选取中间四组(60、70、80、90 cm/s)为实验组。同一规格实验鱼分别在四组流速水平下进行测试, 每次取三尾鱼放入实验装置中, 打开充氧泵, 保持装置上部开放, 将流速水平调至1 BL/s (BL表示体长, 下同), 实验鱼在装置内适应1 h后将流速调至相应的流速水平, 以实验鱼触网达到30 s以上结束, 观测并记录实验鱼的游泳行为和游泳时间, 直至所有规格实验鱼实验完毕。实验过程中, 每隔5 min在探头插孔处用溶氧测定仪测量溶氧水平, 确保DO≥7.00 mg/L, 全程使用摄像机记录实验过程。
1.4.2 Uburst的测定将同一规格实验鱼的三尾鱼放入实验装置中, 将流速水平调至1 BL/s (BL表示体长), 在装置内适应1 h后, 采用逐步增速法测定Uburst。实验初始速度为10 cm/s, 速度增量(∆V)为10 cm/s, 持续游泳时间(T)为20 s, 逐步增加流速水平, 以实验鱼触网达到5 s以上为结束, 观测实验鱼的游泳行为, 直至所有规格实验鱼实验完毕。实验过程中, 保持装置密闭, 每5 min进行游泳装置和水槽的海水交换, 确保密闭游泳装置内的溶氧水平DO≥7.00 mg/L, 实时在探头插孔处用溶氧测定仪测量溶氧水平。Uburst的计算公式如下:
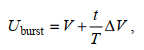 (1)
(1)式中, Uburst为暴发游泳速度(单位: cm/s), V为能够完成设定时间(20 s)的最大游泳速度(单位: cm/s), ∆V为速度增量(10 cm/s), T为设定的持续游泳时间(20 s), t为未能完成设定历时的实际持续游泳时间(t < 20 s)。本实验鱼的鱼体横截面积未超过游泳管截面积的10%, 所以无需校正。
1.4.3 Ucrit的测定将同一规格实验鱼的三尾鱼放入实验装置中, 将流速水平调至1 BL/s, 在装置内适应1 h后, 采用逐步增速法测定Ucrit。实验初始速度为1 BL/s, 速度增量(∆V)为1 BL/s, 持续时间(∆T)为20 min, 逐步增加流速水平, 以实验鱼触网达到20 s以上为结束, 观测实验鱼的游泳行为, 直至所有规格实验鱼实验完毕。实验过程中, 保持装置密闭, 每20 min进行游泳装置和水槽的海水交换, 确保密闭游泳装置内的溶氧水平DO≥7.00 mg/L, 实时在探头插孔处用溶氧测定仪测量溶氧水平。Ucrit的计算公式如下:
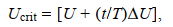 (2)
(2)式中, Ucrit为临界游泳速度(单位: cm/s), U为能够完成设定时间(20 min)的最大游泳速度(单位: cm/s), ∆U为速度增量(1 BL/s), T为设定的持续游泳历时(20 min), t为未能完成设定历时的实际持续游泳时间(t < 20 min)。本实验鱼的鱼体横截面积未超过游泳管截面积的10%, 所以无需校正。
1.4.4 MO2的测定在测定Ucrit的过程中, 每2 min在探头插孔处用溶氧测定仪测定1次实验装置内封闭水体的溶氧值, 记录时间点和溶氧值, 以水体的体积和溶氧值随时间变化斜率的绝对值计算出每尾实验鱼的运动耗氧率MO2。实验开始前, 在实验装置内采用相同的测定方法进行无鱼实验, 得到耗氧细菌和其他生物的耗氧量作为空白对照组。MO2的计算公式如下:
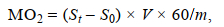 (3)
(3)式中, MO2为运动耗氧率[单位: mg/(kg h)], St为每20 min溶氧值随时间变化的斜率(单位: mgO2/min), S0为空白对照组溶氧值的变化斜率, V为实验装置的容水体积(单位: L), 60为时间常数(单位: min), m为实验鱼体重(单位: kg)。
1.4.5 COT及Uopt的计算COT计算公式如下:
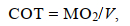 (4)
(4)COT与游泳速度的关系呈幂函数关系, 幂函数关系式为
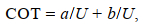 (5)
(5)式中, COT为单位位移耗能[单位: mgO2/(kg m)], MO2为运动耗氧率[单位: mg/(kg h)], V为实验鱼游泳速度(单位: cm/s), U为实验鱼的游泳速度(单位: cm/s)。
将其进行一阶求导并令其等于0, 可求得最适游泳速度, 即单位位移耗能最小时的游泳速度, 计算公式如下:
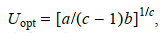 (6)
(6)式中, Uopt为最适游泳速度, a、b和c为常量。
1.5 数据分析实验数据采用SPSS 26进行数据分析。其中每种规格斑石鲷的续航游泳时间、Uburst、Ucrit、MO2、COT、Uopt的差异采用单因素方差分析(One-Way ANOVA); 斑石鲷规格和流速水平对续航游泳时间的影响采用双因素方差分析(Two-way ANOVA); 不同斑石鲷规格的Uburst、Ucrit和Uopt之间的关系采用双变量相关性分析。P < 0.05表示差异性显著, 结果以平均值±标准差(mean±SD)表示。
2 结果 2.1 不同规格斑石鲷的续航游泳时间图 3结果表明, 规格和流速均对四种规格斑石鲷的续航游泳时间有显著影响, 且规格和流速之间存在交互作用(P < 0.001, F=25.401, df=1), 随着规格的增大, 续航游泳时间逐渐增加, 在不同流速水平下350 g斑石鲷的续航游泳时间最长, 且50 g斑石鲷的续航游泳时间在四种流速水平下均显著小于其他三种规格(P < 0.05); 随着流速的增加, 续航游泳时间呈现逐渐减少趋势, 四种规格斑石鲷在60 cm/s流速水平下的续航游泳时间均显著大于在其他三种流速水平下的续航游泳时间(P < 0.05), 续航游泳时间均大于48×102 s, 而在70、80和90 cm/s流速水平下, 50 g和150 g斑石鲷的续航游泳时间均减少至不足6×102 s, 250 g和350 g斑石鲷的续航游泳时间均不足18×102 s, 下降幅度较大。
 |
| 图 3 四种规格斑石鲷在各流速水平下的续航游泳时间 Fig. 3 Endurance swimming time of four specifications of spotted knifejaw at various flow velocity levels 注: 柱上不同字母表示差异显著(P < 0.05) |
随着游泳速度的增加, 四种规格斑石鲷的MO2均呈现上升趋势, 50 g斑石鲷在低于30 cm/s游泳速度时, MO2差异不显著(P > 0.05), 高于30 cm/s游泳速度时, MO2差异显著(P < 0.05); 150 g斑石鲷在低于24 cm/s游泳速度时, MO2差异不显著(P > 0.05), 而高于24 cm/s游泳速度时, MO2差异显著(P < 0.05); 与50 g和150 g斑石鲷MO2曲线变化相比, 250 g和350 g斑石鲷的MO2曲线上升速率更快, 各游泳速度下MO2均显著差异(P < 0.05), 且250 g和350 g斑石鲷MO2的最小值均大于50 g和150 g斑石鲷MO2的最大值(图 4)。
 |
| 图 4 四种规格斑石鲷的MO2 Fig. 4 Exercise oxygen consumption rate of four specifications of spotted knifejaw 注: a. 50 g; b. 150 g; c. 250 g; d. 350 g。图中不同字母表示差异显著(P < 0.05) |
与MO2曲线变化相反, 随着游泳速度的增加, 四种规格斑石鲷的COT均呈现下降趋势(图 5), 逐渐减弱直至曲线趋于平稳。四种规格斑石鲷的COT绝对值分别在初始游泳速度时最高, 且分别在大于40、48、48和63 cm/s游泳速度时, COT差异显著(P < 0.05), 小于40、48、48和63 cm/s游泳速度时, COT无显著差异(P > 0.05)。
 |
| 图 5 四种规格斑石鲷的COT Fig. 5 Energy consumption per unit displacement of four specifications of spotted knifejaw 注: a. 50 g; b. 150 g; c. 250 g; d. 350 g。图中不同字母表示差异显著(P < 0.05) |
图 6结果表明, 随着规格的增大, 四种规格斑石鲷的Ucrit、Uburst和Uopt分别呈现出逐渐升高趋势, 250 g和350 g斑石鲷Uburst均显著大于50 g与150 g (P < 0.05), 50 g和150 g斑石鲷Uburst均无显著差异(P > 0.05); 规格越大, 四种规格斑石鲷的Ucrit呈现出显著升高趋势(P < 0.05), 而与Ucrit变化幅度不同, 虽然Uopt呈现逐渐升高趋势, 但是其变化幅度并不显著(P > 0.05), 规格对四种规格斑石鲷Ucrit的影响均高于Uburst和Uopt。同一规格下斑石鲷Uburst绝对值大于Ucrit, Uopt数值最小, 这表明四种规格斑石鲷的无氧运动能力较强。
 |
| 图 6 四种规格斑石鲷的Ucrit、Uburst和Uopt Fig. 6 Critical swimming speed, burst swimming speed and optimum swimming speed of four specifications of spotted knifejaw 注: 1. Ucrit; 2. Uburst; 3. Uopt。柱上不同字母表示差异显著(P < 0.05) |
图 7结果显示, 50 g斑石鲷Ucrit、Uburst和Uopt两两之间互为负相关关系, 且Uburst和Uopt负相关系数最大, 为–0.77; 150 g斑石鲷Ucrit分别和Uburst、Uopt呈现正相关, 且Ucrit和Uburst相关系数大于Ucrit和Uopt, 而Uburst和Uopt相关性相反, 呈现负相关; 250 g斑石鲷三项游泳能力测定指标之间的关系均不相同, Ucrit和Uburst呈现负相关, Ucrit和Uopt没有相关关系, Uburst和Uopt为正相关关系; 350 g斑石鲷只有Ucrit和Uopt为负相关关系, 且为显著负相关关系(P < 0.05), Uburst分别和Ucrit、Uopt没有相关关系。Uburst为无氧运动能力指标, Ucrit和Uopt为有氧运动能力指标, 通过对四种规格斑石鲷三项游泳能力测定指标进行相关性分析, 结果表明50、150和250 g三种规格斑石鲷的无氧运动能力和有氧运动能力之间均存在关联关系, 均无显著相关性; 350 g斑石鲷无氧运动能力和有氧运动能力之间没有相关关系, 但有氧运动能力中Ucrit和Uopt为显著负相关关系(P < 0.05)。
 |
| 图 7 四种规格斑石鲷Ucrit、Uburst和Uopt之间的关联 Fig. 7 Correlation among critical swimming speed, burst swimming speed and optimum swimming speed of four specifications of spotted knifejaw 注: a. 50 g; b. 150 g; c. 250 g; d. 350 g。A. Ucrit; B. Uburst; C. Uopt。右侧纵坐标表示相关系数 |
续航游泳时间和Uburst反映的是固定流速下的无氧运动能力, 是评价鱼类耐流性能的重要指标(鲜雪梅等, 2010), 续航游泳时间可较全面反映鱼在各种恒定流速条件下的游泳能力, 便于横向比较鱼类之间的游泳能力差异(王萍等, 2010a), Uburst多采用于鱼类在捕食、逃逸、应激反应以及通过鱼道过程中的高流速区域, 对于海水养殖鱼类, 可作为养殖品种抗流能力的重要评价指标(Zeng et al, 2009)。王萍等(2010b)研究认为, 游泳习性是影响续航游泳时间的重要因素, 如果鱼类时常由槽底跃起, 将会增大身体的受力面积, 续航游泳时间减少。本研究结果与这一观点一致, 即实验过程中规格越小的斑石鲷经常由槽底快速向上部做反复游泳运动, 增加了游泳阻力, 续航游泳时间随着规格减少而降低(Lu et al, 2020), 50 g斑石鲷上下游泳运动最为频繁, 且经常做出向前突进游泳动作, 在四种流速下的续航游泳时间均显著小于其他三种规格(P < 0.05)。本研究发现, 在流速为60 cm/s时, 东海网箱养殖主要品种鲈鱼、斜带髭鲷的续航游泳时间分别为25 min和17 min, 四种规格斑石鲷的续航游泳时间均大于80 min, 斑石鲷具有较强的续航游泳能力(王萍等, 2010a)。在70、80和90 cm/s流速下, 50 g和150 g斑石鲷的续航游泳时间减少至不足10 min, 250 g和350 g斑石鲷的续航游泳时间不足30 min, Jones(1971)认为, 在低速时, 鱼类可以不间断地游泳, 耐久游泳时间较高, 对于斑石鲷而言, 60 cm/s及以下为低流速, 高于60 cm/s为高流速。有研究表明, Uburst绝对值随体长的增加而增加, 随着鱼体大小的增加, 鱼体的表面积和肌肉重量的相对值使得绝对游泳速度增大(Videler et al, 1991), 本研究也印证了这一观点, 斑石鲷Uburst随着规格的增加而升高。本研究发现250 g与350 g斑石鲷Uburst显著大于50 g与150 g (P < 0.05), 说明250 g和350 g斑石鲷拥有较强的突进游泳能力, 更适合在深远海进行养殖, 有利于应对突如其来的海上急流, 以免反复被冲击至养殖网箱或围栏网上。与内陆淡水鱼类相比, 生活在江河急流中的长丝裂腹鱼和短须裂腹鱼的平均Uburst分别为124 cm/s和118 cm/s (张沙龙, 2014), 三种鲤科(Phoxinus lagowskii, Triplophysa stoliczkae, Opsariich thysbidens)的平均Uburst为89~113 cm/s (Tan et al, 2021), 相近规格的斑石鲷平均Uburst较高, 为135 cm/s, 我们认为这与斑石鲷生活习性相关, 其生活在海流湍急的近海, 需要高暴发的游泳速度在复杂海况中进行捕食、避敌等游泳动作。
3.2 四种规格斑石鲷的有氧运动能力Ucrit是鱼类有氧游泳能力的上限速度值, 并作为评价不同因素对鱼类游泳能力影响的主要指标, 代表鱼类的最大有氧运动能力(Reidy et al, 2000; Svendsen et al, 2010), 四种规格斑石鲷Ucrit差别显著(P < 0.05), 且绝对值随规格增加而升高(Videler et al, 1991), 规格对斑石鲷Ucrit的影响较为显著, 其原因可能为斑石鲷成长到一定阶段游泳策略发生转变, 如长距离洄游, 繁殖等(Pedersen et al, 2004)。常见于水流较急的新疆特有鱼类斑重唇鱼临界游泳速度为(1.02±0.15) m/s, 同规格斑石鲷Ucrit为61.94~72.34 cm/s, 相比于内陆淡水鱼类斑重唇鱼, 斑石鲷最大有氧运动能力较低(雷青松等, 2020)。通过与续航游泳时间结果对比后发现, 50 g斑石鲷在60 cm/s流速下的续航游泳时间为90 min, 而Ucrit测定实验中60 cm/s流速下持续游泳时间不足20 min, 有研究者认为Ucrit受到实验方案中速度增量、时间间隔带来的累积体力消耗的影响(Pedersen et al, 2004; Hou et al, 2018), 且鱼类多在自然生境采用Ucrit进行游泳运动, 如长距离洄游, 可为斑石鲷游泳能力评价提供参考数据, 但对于深远海养殖参考价值不大(王萍等, 2010a)。
无论持续性游泳能力较强的鱼, 如美国红鱼、中华鲟(Acipenser sinensis)以及大西洋鲑(Salmo salar)等, 还是持续性游泳能力较差的鱼, 如大西洋鳕(Gadus morhua), 其MO2随着速度的增加均呈现上升的趋势(张敬敬等, 2020), 在实验过程中发现, 四种规格斑石鲷与此变化趋势一致, 其MO2随着速度增加而升高(Lee et al, 2003)。在本实验中进一步研究发现, MO2变动幅度存在规格间的差异, 50 g和150 g分别在3倍体长和2倍体长流速之前MO2变化不明显, 表明在此阶段无氧运动占优势; 250 g和350 g变化幅度相同, 各流速下MO2均显著升高, 且数值也明显高于前两种规格, 更早地进入耗氧快速阶段, 即以有氧运动为主(Reidy et al, 2000; Castro-Santos et al, 2012)。以四种规格斑石鲷MO2为基础计算得出COT, 其曲线变化呈现逐渐下降趋势, 有研究结果表明, 生存于不同水流生境鱼类的水流速度选择特征存在明显差异, 与能量效率存在联系, 在同一流速下MO2与COT变化趋势相反, 较低MO2的同时COT较高, 反之亦然(吴青怡等, 2016), 本研究发现四种规格斑石鲷COT均在1倍体长流速水平下数值最大, 在2倍体长流速之后减小直至趋于平缓, 说明在低流速的水平下能量利用率较低, 高流速水平下能量利用率较高(Tudorache et al, 2013)。COT最低时的流速为该规格斑石鲷Uopt, 在工厂化循环水产养殖系统中, 养殖鱼类以Uopt进行游泳运动可以优化其生长性能(Palstra et al, 2015), 且最佳生长的游泳速度很可能接近Uopt, 能量效率最高, 且Uopt在影响鱼类食物摄入的生长方面很重要(Totland et al, 1987; Palstra et al, 2010)。本文认为在斑石鲷养殖过程中, 要根据每种规格Uopt调整循环水流速, 在最佳运动条件下养殖鱼类可以提高生长和食物转化效率, 从而带来更好的经济回报。
3.3 四种规格斑石鲷临界游泳速度、暴发游泳速度和最适游泳速度相关性鱼类的感觉器官受到水流的刺激后, 诱导产生了相应行为和生理反应, 游泳能力之间的关联是探讨生理响应机制的关键(许亚琴, 2020)。王萍等(2010b)认为最大探顶游泳速度、最大续航速度、临界游泳速度在某种程度作为评价鱼类游泳能力的指标具有一致性, Reidy等(2020)以大西洋鳕鱼(Gadus morhua)为实验对象发现, 鱼类有氧耐流运动和无氧暴发运动存在一定关联, 通过对四种规格斑石鲷Uburst、Ucrit和Uopt相关性进行分析, 发现无氧运动能力与有氧运动能力之间存在关联机制, 且结果存在规格上差异(Marras et al, 2010; 曾令清等, 2018)。50 g斑石鲷三项运动能力指标两两之间均为负相关关系, 本研究认为, 斑石鲷成长到50 g时, 其游泳能力发生分化, 处于选择适应生境的运动能力的阶段; 150 g斑石鲷Ucrit均和Uburst、Uopt为正相关关系, Ucrit为鱼类有氧游泳能力的上限速度值, 150 g斑石鲷最大有氧运动能力越高, 其所适应的最佳游泳速度越高, 突进游泳能力也就越高(Palstra et al, 2010), 而Uburst和Uopt呈现负相关, 说明150 g斑石鲷在低能耗的Uopt和高暴发高能耗的Uburst下进行游泳运动存在权衡(Reidy et al, 2000); 250 g斑石鲷游泳能力测定指标之间的相关性与150 g斑石鲷相反, Ucrit和Uburst之间为负相关关系, Uburst和Uopt为正相关关系, 有氧运动能力中Ucrit和Uopt无关联, 说明斑石鲷规格达到250 g时, 其游泳运动能力也随之成长到新的阶段, 低能耗所对应的游泳速度越高, 突进游泳能力也越强(Tan et al, 2021); 斑石鲷规格为350 g时, Ucrit和Uopt为显著负相关关系, Uburst和Uopt、Ucrit均无关联, 本文认为, 在此规格下斑石鲷有氧运动能力之间存在权衡, 会进行最大有氧运动和低能耗有氧运动的选择, 其突进游泳能力处于独立发展阶段, 其运动能力倾向更加明确(Totland et al, 1987; Amelie et al, 2019)。
4 结论综上, 利用工程化围栏或深水网箱等大型深远海养殖设施开展斑石鲷陆海接力养殖过程中要考虑复杂海流、恶劣海况等环境条件, 本研究结果发现: 通过检测不同规格斑石鲷续航游泳时间和Uburst, 将斑石鲷划分为耐流(250 g和350 g)和非耐流(50 g和150 g)两种类型; 斑石鲷放养规格, 应选用250 g及以上且在流速不超过60 cm/s的海区中进行养殖, 如果养殖海域流速过大, 应当采取适当的减流或分流措施, 以保证其最佳流速。上述结果为斑石鲷深远海适宜养殖海域选址和科学放养, 提供了重要理论依据和技术支撑。
致谢 感谢莱州明波水产有限公司在实验过程中提供实验材料和场地,感谢相关部门人员的支持、配合和帮助,谨致谢忱。
王永猛, 柯森繁, 林晨宇, 等, 2021. 红河(元江)流域的典型鱼类游泳能力探究及在过鱼设施流速设计中的应用. 湖泊科学, 33(6): 1820-1831 |
王萍, 桂福坤, 吴常文, 2010a. 美国红鱼(Sciaenops ocellatus)、鲈鱼(Lateolabrax maculatus)、斜带髭鲷(Hapalogenys nitens)耐流性试验研究. 海洋与湖沼, 41(6): 923-929 |
王萍, 桂福坤, 吴常文, 2010b. 鱼类游泳速度分类方法的探讨. 中国水产科学, 17(5): 1137-1146 |
许亚琴, 2020. 流速对拉氏鱥幼鱼生长、非特异性免疫能力及脂肪酸组成的影响[D]. 大连: 大连海洋大学: 11-42.
|
许燕, 王印庚, 张正, 等, 2018. 不同健康程度和抗生素氟苯尼考干预下斑石鲷肠道菌群的结构差异. 水产学报, 42(3): 388-398 |
吴青怡, 曹振东, 付世建, 2016. 鳊鱼和宽鳍鱲幼鱼流速选择与运动能量代谢特征的关联. 生态学报, 36(13): 4187-4194 |
张沙龙, 2014. 长丝裂腹鱼和短须裂腹鱼的游泳能力和游泳行为研究[D]. 武汉: 华中农业大学: 14-35.
|
张敬敬, 朱峰磊, 龙静, 等, 2020. 游泳加速模式对团头鲂和南方鲇的最大游泳速度及运动代谢的影响. 水生生物学报, 44(3): 603-611 |
孟乾, 张志勇, 张志伟, 等, 2020. 斑石鲷和条石鲷线粒体基因组密码子使用分析. 水产科学, 39(5): 702-709 |
赵玉柱, 陈张帆, 马腾, 等, 2021. 斑石鲷性腺发育的组织学观察与初步分析. 上海海洋大学学报, 30(4): 675-683 |
倪海儿, 王国良, 2009. 网箱养殖大黄鱼(Pseudosciaena crocea)疾病与环境因子的关系. 海洋与湖沼, 40(1): 72-77 DOI:10.3321/j.issn:0029-814X.2009.01.012 |
徐琰斐, 徐皓, 刘晃, 等, 2021. 中国深远海养殖发展方式研究. 渔业现代化, 48(1): 9-15 |
徐皓, 刘晃, 徐琰斐, 2021. 我国深远海养殖发展现状与展望. 中国水产, (6): 36-39 |
董登攀, 宋协法, 关长涛, 等, 2010. 褐牙鲆陆海接力养殖试验. 中国海洋大学学报, 40(10): 38-42 |
曾令清, 付成, 付世建, 2018. 鲫幼鱼游泳运动能力的个体变异与表型关联. 水生生物学报, 42(3): 561-570 |
雷青松, 涂志英, 石迅雷, 等, 2020. 应用于鱼道设计的新疆木扎提河斑重唇鱼的游泳能力测试. 水产学报, 44(10): 1718-1727 |
鲜雪梅, 曹振东, 付世建, 2010. 4种幼鱼临界游泳速度和运动耐受时间的比较. 重庆师范大学学报(自然科学版), 27(4): 16-20 |
ARENDT J, HOANG L, 2005. Effect of food level and rearing temperature on burst speed and muscle composition of western spadefoot toad (Spea hammondii). Functional Ecology, 19(6): 982-987 |
AMELIE R, MARC L, CHRISTELLE P, et al, 2019. Relationship between swimming capacities and morphological traits of fish larvae at settlement stage: a study of several coastal Mediterranean species. Journal of Fish Biology, 95(2): 348-356 |
CASTRO-SANTOS T, SANZ-RONDA F J, RUIZ-LEGAZPI J, 2012. Breaking the speed limit-comparative sprinting performance of brook trout (Salvelinus fontinalis) and brown trout (Salmo trutta). Canadian Journal of Fisheries and Aquatic Sciences, 70(2): 280-293 |
FARRELL A P, 2008. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests. Journal of Fish Biology, 72(3): 693-710 |
FU S J, ZENG L Q, LI X M, et al, 2009. The behavioural, digestive and metabolic characteristics of fishes with different foraging strategies. Journal of Experimental Biology, 212(14): 2296-2302 |
HOOVER J J, ZIELINSKI D P, SORENSEN P W, 2017. Swimming performance of adult bighead carp Hypophthalmichthys nobilis (Richardson.1845) and silver carp H. molitrix (Valenciennes.1844). Journal of Applied Ichthyology, 33(1): 54-62 |
HOU Y, CAI L, WANG X, et al, 2018. Swimming performance of 12 Schizothoracinae species from five rivers. Journal of Fish Biology, 92(6): 2022-2028 |
JIA Y D, GAO Y T, GAO Y H, et al, 2021. Growth performance, hematological and biochemical parameters, and hepatic antioxidant status of spotted knifejaw Oplegnathus punctatus in an offshore aquaculture net pen. Aquaculture, 541: 736761 |
JOHANSSON D, LAURSEN F, FERNÖ A, et al, 2014. The interaction between water currents and salmon swimming behaviour in sea cages. PLoS One, 9(5): e97635 |
JONES D R, 1971. The effect of hypoxia and anaemia on the swimming performance of rainbow trout (Salmo gairdneri). Journal of Experimental Biology, 55(2): 541-551 |
KAUFMANN R, WIESER W, 1992. Influence of temperature and ambient oxygen on the swimming energetics of cyprinid larvae and juveniles. Environmental Biology of Fishes, 33(1/2): 87-95 |
LEE C G, FARRELL A P, LOTTO A, et al, 2003. The effect of temperature on swimming performance and oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O. kisutch) salmon stocks. Journal of Experimental Biology, 206(18): 3239-3251 |
LU C, CHEN J H, JOHNSON D, et al, 2020. Effect of body length on swimming capability and vertical slot fishway design. Global Ecology and Conservation, 22: e00990 |
MEHNER T, WIESER W, 1994. Energetics and metabolic correlates of starvation in juvenile perch (Perca fluviatilis). Journal of Fish Biology, 45(2): 325-333 |
MARRAS S, CLAIREAUX G, MCKENZIE D, et al, 2010. Individual variation and repeatability in aerobic and anaerobic swimming performance of European sea bass, Dicentrarchus labrax. The Journal of Experimental Biology, 213(1): 26-32 |
NOBATA S, HOUKI S, KITAGAWA T, et al, 2022. Condition factor dependency of burst swimming ability between wild and hatchery-reared chum salmon fry (Oncorhynchus keta). Ichthyological Research, 69(2): 280-286 |
PALSTRA A P, MES D, KUSTERS K, et al, 2015. Forced sustained swimming exercise at optimal speed enhances growth of juvenile yellowtail kingfish (Seriola lalandi). Frontiers in Physiology, 5: 506 |
PALSTRA A P, TUDORACHE C, ROVIRA M, et al, 2010. Establishing zebrafish as a novel exercise model: swimming economy, swimming-enhanced growth and muscle growth marker gene expression. PLoS One, 5(12): e14483 |
PEDERSEN L F, MALTE H, 2004. Repetitive acceleration swimming performance of brown trout in fresh water and after acute seawater exposure. Journal of Fish Biology, 64(1): 273-278 |
PETERSON R H, ANDERSON J M, 1969. Influence of temperature change on spontaneous locomotor activity and oxygen consumption of Atlantic salmon, Salmo salar, acclimated to two temperatures. Journal of the Fisheries Board of Canada, 26(1): 93-109 |
PLAUT I, 2000. Effects of fin size on swimming performance, swimming behaviour and routine activity of zebrafish Danio rerio. Journal of Experimental Biology, 203(4): 813-820 |
REIDY S P, KERR S R, NELSON J A, 2000. Aerobic and anaerobic swimming performance of individual Atlantic cod. Journal of Experimental Biology, 203(2): 347-357 |
RICHER E E, FETHERMAN E R, KRONE E A, et al, 2020. Multispecies fish passage evaluation at a rock-ramp fishway in a Colorado transition zone stream. North American Journal of Fisheries Management, 40(6): 1510-1522 |
SVENDSEN J C, TUDORACHE C, JORDAN A D, et al, 2010. Partition of aerobic and anaerobic swimming costs related to gait transitions in a labriform swimmer. Journal of Experimental Biology, 213(13): 2177-2183 |
TAN J J, TAN H L, GOERIG E, et al, 2021. Optimization of fishway attraction flow based on endemic fish swimming performance and hydraulics. Ecological Engineering, 170: 106332 |
TUDORACHE C, DE BOECK G, CLAIREAUX G, 2013. Forced and preferred swimming speeds of fish: a methodological approach[M]//PALSTRA A P, PLANAS J V. Swimming Physiology of Fish. Berlin, Germany: Springer: 81-108.
|
TOTLAND G K, KRYYI H, JØDESTØL K, et al, 1987. Growth and composition of the swimming muscle of adult Atlantic salmon (Salmo salar L.) during long-term sustained swimming. Aquaculture, 66(3/4): 99-313 |
VIDELER J J, WARDLE C S, 1991. Fish swimming stride by stride: speed limits and endurance. Reviews in Fish Biology and Fisheries, 1(1): 23-40 |
WANG J T, HUANG R X, HAN T, et al, 2021. Dietary protein requirement of juvenile spotted knifejaw Oplegnathus punctatus. Aquaculture Reports, 21: 100874 |
WATKINS T B, 1996. Predator-mediated selection on burst swimming performance in tadpoles of the Pacific tree frog, Pseudacris regilla. Physiological Zoology, 69(1): 154-167 |
WESTERTERP K, 1977. How rats economize: energy loss in starvation. Physiological Zoology, 50(4): 331-362 |
XU W T, LI M, WANG S, et al, 2022. Transcriptomic profiles of spotted knifejaw (Oplegnathus punctatus) spleen in response to Megalocytivirus infection. Aquaculture, 555: 738212 |
YANASE K, EAYRS S, ARIMOTO T, 2007. Influence of water temperature and fish length on the maximum swimming speed of sand flathead, Platycephalus bassensis: implications for trawl selectivity. Fisheries Research, 84(2): 180-188 |
YLIEFF M Y, PONCIN P, 2003. Quantifying spontaneous swimming activity in fish with a computerized color video tracking system, a laboratory device using last imaging techniques. Fish Physiology and Biochemistry, 28(1/2/3/4): 281-282 |
ZENG L Q, CAO Z D, FU S J, et al, 2009. Effect of temperature on swimming performance in juvenile southern catfish (Silurus meridionalis). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 153(2): 125-130 |