 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 卢占晖, 朱文斌, 徐开达, 钟志, 周永东. 2023.
- LU Zhan-Hui, ZHU Wen-Bin, XU Kai-Da, ZHONG Zhi, ZHOU Yong-Dong. 2023.
- 浙江沿岸海域螺贝类群落结构及其季节变化
- STUDY ON SEASONAL CHANGES OF SNAILS AND SHELLFISH COMMUNITIES IN COSTAL AREAS OFF ZHEJIANG PROVINCE
- 海洋与湖沼, 54(1): 276-285
- Oceanologia et Limnologia Sinica, 54(1): 276-285.
- http://dx.doi.org/10.11693/hyhz20220400108
文章历史
-
收稿日期:2022-04-26
收修改稿日期:2022-07-19




浙江沿岸海域受长江冲淡水、台湾暖流、苏北沿岸水和黄海冷水团等海流和水系的综合影响, 加之沿岸海域岛屿分布众多, 各水系交流充分, 营养盐类和饵料生物丰富, 是多种包括螺贝类在内的海洋生物的产卵场和索饵场(何贤保等, 2013)。螺贝类作为软体动物门中最重要的类群, 广泛栖息于海洋、淡水以及陆地等各种环境中。在海洋生态系统中, 它们既可作为分解者分解动植物残体和有机碎屑, 又能作为消费者牧食大型藻类和维管植物, 还能作为次级生产者被鱼类和甲壳类等多种海洋动物所摄食(Brönmark, 1990; Elger et al, 2005)。因此, 螺贝类是海洋生态系统物质循环和能量流动的重要环节之一; 另外, 螺贝类等大型底栖动物的群落组成与结构对自然环境变化和人类干扰极为敏感, 某些种类(如光滑河篮蛤Potamocorbula laevis, 缢蛏Sinonovacula constricta)是监测海洋生态环境变化的良好指示生物(王金宝等, 2018)。目前, 关于浙江沿岸螺贝类种类组成、分布特征、资源密度以及生态特征等方面的研究, 主要集中在岛礁区的潮间带海域(杨俊毅等, 2000; 邹莉等, 2010; 蔡林婷等, 2013; 陈万东等, 2017), 或者仅作为近海大型底栖生物群落的重要组成类群之一进行研究(周细平等, 2019; 梁静香等, 2020; 王航俊等, 2020; 严润玄等, 2020), 而对于近海海域螺贝类群落结构方面的专门研究尚未见报道。
本研究基于2019年全年4个航次浙江近岸海域渔业资源大面定点调查数据, 研究分析了螺贝类群落的种类组成、优势种组成、资源密度分布、群落物种多样性以及外部因素对其群落结构的干扰程度等内容, 系统阐述了沿岸螺贝类群落结构特征及其季节变化。此研究的开展, 有助于丰富中国近海海域软体动物生物多样性的基础资料, 对于认识浙江乃至整个东海海域螺贝类最新的资源动态变化有一定的科学意义, 同时也为浙江近海海洋生态和环境保护措施的制定提供参考依据。
1 材料与方法 1.1 数据来源本文所用数据来自2019年1月(冬季)、4月(春季)、7月(夏季)和11月(秋季)进行的“浙江渔场渔业资源动态监测调查”课题。共设置调查站位87个, 站位分布如图 1所示。调查船为单船底拖网渔船, 船长38 m, 总吨位150 t, 主机功率202 kW。调查网具网口拉紧周长50 m, 囊网网目尺寸25 mm, 上纲长30 m, 下纲长38 m。各站位拖网时间均为1 h, 平均拖速3 kn。各站位渔获物样品随机采样1箱, 渔获物中螺贝类种类尽可能鉴定到最小分类单元, 并记录每种的生物量和尾数, 螺贝类种类名称以《中国海洋生物名录》(刘瑞玉, 2008)为依据, 同时参照Sealifebase数据库(http://www.sealifebase.org)。上述调查采样及测定均按《海洋调查规范(GB/T 12763.6-2007)》(中华人民共和国国家质量监督检验检疫总局等, 2008)的有关标准进行。根据渔场环境条件及其生物种类分布特征, 将调查海域分为3个区域: 即将29°30′N以北(1~38号站位)水域称为北部海域, 28°30′N以南(63~87号站位)水域称为南部海域, 28°30′~29°30′N (39~62号站位)之间的水域称为中部海域。
 |
| 图 1 浙江沿岸海域调查站位分布图 Fig. 1 Location of sampling stations in coastal areas of Zhejiang Province 注: 北部: 1~38号; 中部: 39~62号; 南部: 63~87号 |
群落优势种与常见种采用Pinkas的相对重要性指数(index of relative importance, IRI)进行划分(Pinkas et al, 1971)。其公式为
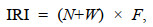 (1)
(1)式中, N为某一种类的尾数占总尾数的百分比(单位: %); W为某一种类的生物量占总生物量的百分比(单位: %); F为某一种类出现的站数占调查总站数的百分比(单位: %)。根据不同螺贝类优势度(IRI)的大小, 确定不同种类在群落内的重要性。本研究将相对重要性指数(IRI)大于1 000的种类定为优势种, 500~1 000之间的种类定为常见种(袁伟等, 2020)。
1.2.2 资源密度螺贝类主要营底栖生活, 因此, 本研究采用扫海面积法对调查海域螺贝类资源密度分布进行分析。其计算公式为
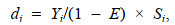 (2)
(2)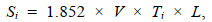 (3)
(3)式中, di为i站位的资源密度(单位: kg/km2), Yi为调查船在i站位的渔获量(单位: kg); Si为调查船在i站位的扫海面积(单位: km2); E为逃逸率(根据本次调查网具——单船底拖网的结构, 渔法特点以及螺贝类分布水层和生活习性, 综合相关研究报告确定逃逸率E的取值) (Aglen et al, 1981; 卢占晖等, 2018), 根据以上原则本研究拟选取的逃逸率为0.2; V为网具拖曳的平均拖速(单位: kn) (本次调查拖网拖曳平均拖速为3 kn); Ti为拖网时间(本次调查均为1 h); L为网口水平扩张宽度(单位: km) (本次调查网具网口宽度为5×10–3 km)。
1.2.3 群落多样性本文螺贝类群落多样性分析运用下列公式(Margalef, 1958):
种类丰富度指数D:
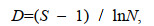 (4)
(4)Shannon-Wiener多样度指数H′:
 (5)
(5)种类均匀度指数J′:
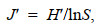 (6)
(6)式中, S为某站位的螺贝类种类数; N为该站位总个体数; Pi为该站位第i种螺贝类个体数占螺贝类总渔获个体数的比例。
1.2.4 群落ABC曲线本研究根据ABC曲线中生物量和丰度的K-优势度曲线, 分析检验浙江沿岸海域螺贝类群落受到污染和扰动的情况。若生物量曲线位于丰度曲线之上, 则表明群落处于未受干扰(稳
定)的状态; 若两条曲线相交, 则表明群落处于中等干扰(或不稳定)的状态; 若生物量曲线在丰度曲线之下, 则表明群落处于严重干扰的(不稳定的)状态。用W统计量(W-statistic)作为ABC曲线方法的一个统计量, 其公式为:
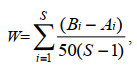 (7)
(7)式中, Bi和Ai为ABC曲线中种类序号对应的生物量和丰度的累积百分比, S为出现物种数。当生物量优势曲线在丰度优势度曲线之上时, W为正, 反之则W为负(Clarke et al, 2001)。
以上数据分析中, 群落的物种多样性指数、ABC曲线及W统计量的计算均使用PRIMER6.0软件进行处理; 站位图、种类空间分布和资源密度分布图利用Surfer8.0作图软件进行绘制。
2 结果 2.1 种类组成与分布本次全年4个季节的调查中共采集螺贝类43种, 隶属于2纲, 7目, 26科。其中, 腹足纲(螺类)出现种类30种, 占螺贝类总种类数的69.77%, 双壳纲(贝类)出现种类13种, 占螺贝类总种类数的30.23%; 新腹足目出现种类最多, 有17种, 占螺贝类总种类数的32.56%, 其次为中腹足目、帘蛤目和蚶目, 种类数分别为15种(34.88%)、6种(13.95%)和2种(4.65%), 其余异腹足目、珍珠贝目和贻贝目出现的种类均为1种(表 1)。
| 纲 | 目 | 科 | 种 | 季节 | |||
| 春 | 夏 | 秋 | 冬 | ||||
| 腹足纲Gastropoda | 新腹足目Neogastropoda | 蛾螺科Buccinidae | 褐管蛾螺Siphonalia spadicea | + | + | + | + |
| 泥东风螺Babylonia lutosa | + | ||||||
| 骨螺科Muricidae | 红螺Rapana bezoar | + | |||||
| 脉红螺Rapana venosa | + | + | + | + | |||
| 浅缝骨螺Murex trapa | + | + | + | + | |||
| 衲螺科Cancellariidae | 金刚衲螺Cancellaria spengleriana | + | + | + | + | ||
| 笋螺科Terebridae | 珍笋螺Terebra pretiosa | + | |||||
| 塔螺科Pyramidellidae | 爪哇拟塔螺Turricula javana | + | + | + | |||
| 白龙骨乐飞螺Lophiotomo leucotropis | + | + | |||||
| 假奈拟塔螺Turricula nelliaespurius | + | + | + | ||||
| 洁裁判螺Inquisitor recurvirostrata | + | + | + | + | |||
| 细带螺科Fasciolariidae | 长尾纺锤螺Fusinus longicaudus | + | + | + | + | ||
| 织纹螺科Nassariidae | 纵肋织纹螺Nassarius variciferus | + | + | + | |||
| 细肋织纹螺Nassarius castus | + | + | + | ||||
| 红带织纹螺Nassarius succinctus | + | + | |||||
| 方格织纹螺Nassarius conoidalis | + | ||||||
| 西格织纹螺Nassarius siquijorensis | + | + | + | ||||
| 中腹足目Mesogastropoda | 鹑螺科Doliidae | 斑鹑螺Tonna dolium | + | + | + | ||
| 带鹑螺Tonna olearium | + | + | + | + | |||
| 沟鹑螺Tonna sulcosa | + | + | + | ||||
| 中国鹑螺Tonna chinensis | + | ||||||
| 冠螺科Cassididae | 双沟鬘螺Phalium bisulcatum | + | + | + | + | ||
| 盔螺科Galeodidae | 管角螺Hemifusus tuba | + | + | + | + | ||
| 角螺Hemifusus colosseue | + | ||||||
| 琵琶螺科Ficidae | 长琵琶螺Ficus gracilis | + | |||||
| 梭螺科Ovulidae | 钝梭螺Volva volva | + | |||||
| 蛙螺科Bursidae | 习见蛙螺Bursa rana | + | + | + | + | ||
| 涡螺科Volutidae | 电光螺Fulgoraria rupestrisrupestris | + | + | + | + | ||
| 衣笠螺科Xenophoridae | 光衣笠螺Xenophora exuta | + | + | + | + | ||
| 玉螺科Naticidae | 扁玉螺Nevetita didyma | + | + | + | + | ||
| 爪哇窦螺Sinum javanicum | + | + | + | ||||
| 锥螺科Turritellidae | 棒锥螺Turritella bacillum | + | + | + | + | ||
| 异腹足目Heterogastropoda | 光螺科Eulimidae | 马氏光螺Melanella martinii | + | ||||
| 双壳纲Bivalvia | 帘蛤目Veneroida | 樱蛤科Tellinidae | 彩虹明樱蛤Moerella iridescens | + | + | ||
| 美女白樱蛤Macoma candida | + | + | + | + | |||
| 刀蛏科Pharidae | 小刀蛏Cultellus attenuatus | + | + | + | |||
| 帘蛤科Veneridae | 条纹卵蛤Costellipitar chordatum | + | |||||
| 刀蛏科Cultellidae | 小荚蛏Siliqua milimai | + | + | ||||
| 截蛏科Solecurtidae | 缢蛏Sinonovacula constricta | + | |||||
| 蚶目Arcoida | 蚶科Arcidae | 毛蚶Scapharca kagoshimensis | + | + | + | + | |
| 泥蚶Tegillarca granosa | + | + | + | + | |||
| 珍珠贝目Pterioida | 牡蛎科Ostreidae | 近江牡蛎Ostrea rivularis | + | ||||
| 贻贝目Mytiloida | 贻贝科Mytilidae | 紫贻贝Mytilus edulis | + | ||||
| 注: + 表示在该季节出现 | |||||||
从种类组成的季节分布看, 春季: 该调查航次出现的螺贝类种类为32种, 隶属于2纲, 5目, 19科, 为全年4个季节中出现种类数最多的季节。该航次调查的87个站位中, 出现螺贝类的调查站位为70个, 出现频率为80.46%, 其不同站位间出现的螺贝类种类数最多为8种, 出现在调查海域中北部区域的5、32和39号站位。春季螺贝类种类数的分布呈现由北至南逐渐减少的趋势, 空间分布特征较为明显(表 1, 图 2)。
 |
| 图 2 浙江沿岸海域螺贝类种类空间分布 Fig. 2 The space distribution of snails and shellfish species in coastal areas of Zhejiang Province |
夏季: 该调查航次出现的螺贝类种类为27种, 隶属于2纲, 4目, 18科, 为全年4个季节中出现种类数最少的季节。该航次调查的87个站位中, 出现螺贝类的调查站位为55个, 出现频率为63.22%, 其不同站位间出现的螺贝类种类数最多为5种, 出现在调查海域中部区域的58号站位。夏季螺贝类种类数的分布特征与春季相反, 呈现由北至南逐渐增加的趋势, 空间分布特征较为明显(表 1, 图 2)。
秋季: 该调查航次出现的螺贝类种类为28种, 隶属于2纲, 6目, 20科。该航次调查的87个站位中, 出现螺贝类的调查站位为54个, 出现频率为62.07%, 其不同站位间出现的螺贝类种类数最多为6种, 分别出现在调查海域北部区域的21号和南部区域的82号站位。秋季不同站位出现螺贝类种类数分布较为均匀, 仅在调查海域北部和南部的个别站位出现种类数相对较多, 空间分布特征不明显(表 1, 图 2)。
冬季: 该调查航次出现的螺贝类种类为29种, 隶属于2纲, 4目, 20科。该航次调查的87个站位中, 出现螺贝类的调查站位为66个, 出现频率为75.86%, 其不同站位间出现的螺贝类种类数最多为10种, 出现在调查海域北部区域的5号站位。冬季不同站位出现螺贝类种类数的高值区主要集中在调查海域的北部外侧区域, 即调查海域螺贝类种类数的空间分布呈现北部高, 中南部低的趋势(表 1, 图 2)。
2.2 优势种与常见种利用相对重要性指数(IRI)来确定各种类在螺贝类群落中的重要性(表 2)。根据“材料与方法”中的规定: 春季螺贝类群落的优势种按IRI值大小依次为褐管蛾螺、棒锥螺和习见蛙螺, 这3种螺贝类渔获质量占该航次螺贝类总渔获物质量的71.80%, 渔获尾数占螺贝类总渔获物尾数的78.92%, 春季螺贝类群落无常见种; 夏季螺贝类群落的优势种为棒锥螺, 该种类渔获质量占该航次螺贝类总渔获物质量的58.45%, 渔获尾数占螺贝类总渔获物尾数的65.87%, 常见种为褐管蛾螺; 秋季螺贝类群落的优势种按IRI值大小依次为棒锥螺和习见蛙螺, 2个种类渔获质量占该航次螺贝类总渔获物质量的75.72%, 渔获尾数占螺贝类总渔获物尾数的83.22%, 秋季螺贝类群落无常见种; 冬季螺贝类群落的优势种为棒锥螺, 该种类渔获质量占该航次螺贝类总渔获物质量的41.80%, 渔获尾数占螺贝类总渔获物尾数的44.69%, 常见种为长尾纺锤螺和扁玉螺。
| 季节 | 种类 | F/% | N/% | W/% | IRI |
| 春 | 褐管蛾螺 | 24.14 | 40.22 | 23.59 | 1 540.23 |
| 棒锥螺 | 21.84 | 31.94 | 29.58 | 1 343.45 | |
| 习见蛙螺 | 41.38 | 6.76 | 18.64 | 1 051.00 | |
| 夏 | 棒锥螺 | 18.39 | 65.87 | 58.45 | 2 286.25 |
| 褐管蛾螺 | 13.79 | 21.29 | 15.08 | 501.64 | |
| 秋 | 棒锥螺 | 12.64 | 66.89 | 48.87 | 1 463.60 |
| 习见蛙螺 | 32.18 | 16.33 | 26.85 | 1 389.48 | |
| 冬 | 棒锥螺 | 21.84 | 44.69 | 41.80 | 1 888.99 |
| 长尾纺锤螺 | 17.24 | 17.70 | 22.39 | 691.18 | |
| 扁玉螺 | 24.14 | 12.81 | 8.70 | 519.05 | |
| 注: W表示占总生物量的百分比; N表示占总尾数的百分比; F表示出现频率; IRI表示相对重要性指数 | |||||
在调查海域的螺贝类群落中, 全年4个季节均出现的优势种为棒锥螺, 且除春季以外, 棒锥螺均为群落中的第一优势种, 因此棒锥螺为浙江沿岸螺贝类群落中的绝对优势种。其次, 习见蛙螺为春、秋季两个季节的优势种, 而褐管蛾螺则为春季调查航次中螺贝类群落中的第一优势种。总体来看: 调查海域全年螺贝类群落的优势种与常见种的种类数共有5种, 且优势种种类组成的季节变化不大。
2.3 资源密度全年4个航次调查中共采集螺贝类52 194个, 生物量574.02 kg, 平均个体生物量11.00 g, 4个航次调查总站位数为348个, 其中出现螺贝类的站位数有245个, 出现频率70.40%。4个航次调查海域螺贝类平均资源密度的大小为夏季 > 秋季 > 春季 > 冬季。
春季: 螺贝类平均资源密度为158.30 kg/km2, 资源密度最小值为0.19 kg/km2, 出现在位于中部海域的44号站位, 资源密度最大值为1 276.94 kg/km2, 出现在位于中部海域的47号站位; 根据不同站位间资源密度的空间分布状况(图 3), 整个调查海域螺贝类资源密度的高值区主要集中在调查海域的中南部区域, 即29°00′N以南海域, 且外侧调查站位的资源密度明显高于内侧调查站位, 空间分布特征较为明显。
 |
| 图 3 浙江沿岸海域螺贝类资源密度的季节分布 Fig. 3 The seasonal distribution in resource density of snails and shellfish in coastal areas of Zhejiang Province |
夏季: 螺贝类平均资源密度为356.27 kg/km2, 为全年最高值, 资源密度最小值为2.40 kg/km2, 出现在位于北部海域的26号站位, 资源密度最大值为7 225.40 kg/km2, 出现在位于南部海域的69号站位; 根据不同站位间资源密度的空间分布状况(图 3), 整个调查海域螺贝类资源密度的高值区主要集中在调查海域的南部区域, 即28°30′N以南海域, 空间分布特征较为明显。
秋季: 螺贝类平均资源密度为200.85 kg/km2, 为全年次高值, 资源密度最小值为0.43 kg/km2, 出现在位于北部海域的11号站位, 资源密度最大值为3 939.74 kg/km2, 出现在位于南部海域的84号站位; 根据不同站位间资源密度的空间分布状况(图 3), 整个调查海域螺贝类资源密度的高值区主要集中在调查海域的南部区域, 即28°00′N以南海域, 空间分布特征较为明显。
冬季: 螺贝类平均资源密度为132.82 kg/km2, 为全年最低值, 资源密度最小值为0.14 kg/km2, 出现在位于北部海域的15号站位, 资源密度最大值为2 126.86 kg/km2, 出现在位于南部海域的69号站位; 根据不同站位间资源密度的空间分布状况(图 3), 整个调查海域螺贝类资源密度的呈现中间低, 南北高的空间分布趋势。
2.4 物种多样性选取3个具有代表性的生物多样性特征值: 种类丰富度指数(D)、以生物量计算的Shannon-Wiener多样度指数(H′)和以生物量计算的种类均匀度指数(J′)来分析浙江沿岸海域螺贝类群落多样性特点(表 3)。经计算可得: 春季3种多样性指数的平均值分别为0.68 (0~1.76)、0.74 (0~1.54)和0.64 (0~0.98); 夏季3种多样性指数的平均值分别为0.43 (0~0.84)、0.65 (0~1.39)和0.66 (0~0.99); 秋季3种多样性指数的平均值分别为0.43 (0~1.25)、0.64 (0~1.60)和0.67 (0~1.00);冬季3种多样性指数的平均值分别为0.69 (0~1.49)、0.82 (0~1.91)和0.65 (0~0.98)。全年4个季节3种群落多样性指数数值均较小, 螺贝类群落多样性水平处于较低水平。单因素方差分析表明, 浙江沿岸海域螺贝类群落结构多样性指数全年差异均不显著(P > 0.05)。
| 季节 | 物种丰富度指数(D) | 物种多样性指数(H′) | 均匀度指数(J) | |||||
| 均值 | 范围 | 均值 | 范围 | 均值 | 范围 | |||
| 春 | 0.68±0.01 | 0~1.76 | 0.74±0.01 | 0~1.54 | 0.64±0.01 | 0~0.98 | ||
| 夏 | 0.43±0.02 | 0~0.84 | 0.65±0.01 | 0~1.39 | 0.66±0.01 | 0~0.99 | ||
| 秋 | 0.43±0.02 | 0~1.25 | 0.64±0.04 | 0~1.60 | 0.67±0.03 | 0~1.00 | ||
| 冬 | 0.69±0.02 | 0~1.49 | 0.82±0.01 | 0~1.91 | 0.65±0.03 | 0~0.98 | ||
4个季节的ABC曲线的趋势如图 4所示。所有4个季节螺贝类群落的丰度优势度曲线与生物量的优势度曲线均相交。4个季节根据ABC曲线计算的W统计值变化范围为–0.035~0.007。其中, 春、夏和秋季三个季节的W统计值均为负数, 冬季W统计值为正数。根据Warwick和Clarke的理论, 4个季节螺贝类群落均处于中等干扰(或不稳定)的状态(Clarke et al, 2001), 但冬季螺贝类群落受到的外界干扰相对最低。
 |
| 图 4 浙江沿岸海域螺贝类群落的ABC曲线以及W统计值 Fig. 4 ABC curves and W value of snails and shellfish community in coastal areas of Zhejiang Province 注: ▽表示生物量; Δ表示丰度 |
本次全年4个季节的调查中共采集螺贝类43种, 隶属于7目, 26科。与浙江沿岸(26种)、舟山东极(33种)以及玉环披山(33种)等岛礁区(邹莉等, 2010; 蔡林婷等, 2013; 陈万东等, 2017)等历史调查资料相比较, 本次调查所捕获的种类数均高于以往历史数据, 而略低于浙江动物志中该区域记录种类58种(蔡如星等, 2001), 但本次调查捕获的螺贝类基本涵盖了浙江近海海域螺贝类的主要种类。主要是由于本次调查的范围更为广泛, 捕获外海种类的概率更高。同其他邻近海区相比, 本次调查出现的种类数高于山东近海海域种类(15种) (赵凡淇等, 2018)和南黄海近海岛礁海域(34种) (祝超文等, 2022), 但显著低于闽江口岛礁区海域(61种) (林建杰, 2022)和南海三亚岛礁区海域(62种) (马文刚等, 2022), 这也能够从一定程度上说明热带海域较亚热带和温带海域具有更多螺贝类种类。从种类数的空间分布特征来看, 全年4个航次调查海域种类分布的特征均呈现出由北向南种类数逐渐增多的趋势, 这主要是受到外海黑潮暖流以及水温逐渐增高的影响, 南部海域出现了更为典型的热带性种类(如管角螺、沟鹑螺等), 这一结果与以前的研究资料也较为一致(蔡如星等, 2001)。浙江沿岸海域螺贝类群落共出现3种优势种, 分别为棒锥螺、褐管蛾螺和习见蛙螺, 以上3种种类均属于高温广盐类群, 这也说明浙江沿岸海域螺贝类群落以该类群占主导地位。棒锥螺是4个季节的共有优势种, 且优势度明显高于其他两个优势种以及群落中的其他种类, 说明研究海域螺贝类优势种较为稳定, 群落的性质和功能可通过少数种类进行控制。这一点在其他大型底栖动物群落的结构特征中同样得以证实(刘迅等, 2017)。根据调查结果, 浙江沿岸4个季节群落多样性指数季节间变化不明显, 且整体水平偏低, 群落多样性水平不高, 群落处于相对不稳定状态。3个群落多样性水平指数的空间分布特征均呈现沿岸低, 外海高的趋势。
3.2 资源密度分布特征浙江沿岸海域螺贝类资源密度存在明显的季节变化。其中, 夏季最高, 秋季次之, 冬季最低。究其原因, 其一是由于影响螺贝类群落资源密度的高低的种类——棒锥螺、褐管蛾螺和习见蛙螺(三种种类对全年4个航次螺贝类的平均资源密度贡献率之和均超过70%), 而以上3种种类均属于暖水性种类, 其资源密度的高低与海水温度尤其是底层温度层显著的正相关, 即水温越高, 资源密度越高, 海水温度高低的分布也恰好为夏季最高, 冬季最低; 其二是因为夏季航次恰好处于“伏季休渔”期间, 调查海域捕捞活动的影响为全年最小, 其资源养护效果最为明显, 平均资源密度则相对最高, 而随着休渔期结束后秋冬汛的来临, 捕捞强度加大, 则资源密度逐渐降低。其资源密度夏高冬低的季节变化在浙江沿岸其他底栖动物群落的研究结果中同样得以印证(陈伟峰等, 2018; 丁朋朋等, 2019)。根据图 3所体现的资源密度空间分布趋势可得: 浙江近岸海域螺贝类资源密度总体上呈现由北向南逐渐增高的趋势, 此现象与以往浙江沿岸贝类的分布特征保持高度一致(邹莉等, 2010)。需要特别指出的一点: 以往的研究多采用阿氏拖网或者采泥器进行样品取样, 前者属于定性/半定量采集网具, 最小网囊网目尺寸25 mm, 网口宽度1.5 m; 后者虽为定量采样工具, 但其采样面积仅为0.1 m2, 捕获活动能力强的种类的概率与效率均相对较差。而本次采样使用的采样网具无论从采样的覆盖范围、机动能力均强于上述两种网具, 因而此次调查可以更为准确地反映研究区域螺贝类资源密度的时空分布特征。
3.3 扰动对群落的影响ABC曲线已经被广泛应用于监测干扰对包括螺贝类在内的海洋底栖无脊椎动物群落的影响(李少文等, 2017; 金文育等, 2017; 胡桂坤等, 2019)。综合调查海域全年螺贝类群落ABC曲线特征及W统计值, 根据Clarke和Warwick的划分标准, 表明调查海域螺贝类群落全年均处于中等干扰(或不稳定)的状态, 而此时群落中生物量占优势的大个体消失, 在数量上占优势的种是个体较小的种, 在此情况下种内丰度的分布与生物量分布优势难分(Clarke et al, 2001)。Warwick(1986)根据调查中出现的螺贝类个体大小, 平均体重50 g以上的大型螺贝类种类(脉红螺、沟鹑螺等)的生物量仅占总生物量的9.94%, 而平均体重在10 g以下的小型种类(棒锥螺、泥蚶等)数量则占总数量的86.11%, 充分验证了螺贝类群落于中等干扰状态这一论断。究其原因, 主要是由于底质的稳定性是衡量其是否适宜包括螺类在内的底栖动物生存的基本条件之一, 底质任何形式的不稳定均可对包括螺贝类在内的大型底栖生物产生不利影响(李亚芳等, 2016), 而近年来在浙江近海渔场从事捕捞活动且对底质破坏较为严重的拖网(单拖、双拖以及桁杆拖虾)作业日益频繁, 加之不断增长的涉海工程、海洋污染等人为因素对底栖生境的破坏, 对螺贝类群落的生境构成了较为严重的影响。需要指出的是: 包括螺贝
类在内的大型底栖生物群落, 其结构受多种环境因子综合作用的影响, 但由于调查项目的局限性, 诸如温盐深、营养盐、悬浮物、pH、沉积物粒径、浮游动植物丰度等生物和非生物环境因子对螺贝类种类分布与群落结构的影响未能涉及, 这也有待于进一步研究。
4 结论(1) 种类组成与优势种浙江沿岸海域全年4个季节调查中共采集螺贝类43种, 隶属于2纲, 7目, 26科。春季出现种类最多, 夏季出现种类最少。从种类数的空间分布特征来看, 全年4个航次调查海域种类分布的特征均呈现出由北向南种类数逐渐增多的趋势。棒锥螺为浙江沿岸螺贝类群落中的绝对优势种, 优势种种类组成的季节变化不大。
(2) 资源密度时空分布浙江沿岸海域螺贝类资源密度存在明显的季节变化。其中, 夏季最高, 秋季次之, 冬季最低。浙江沿岸海域螺贝类资源密度空间分布总体上呈现由北向南逐渐增高的趋势, 此现象与以往浙江沿岸贝类的分布特征保持高度一致。
(3) 群落的动态变化根据ABC曲线特征及W统计值, 研究海域螺贝类群落全年均处于中等干扰(或不稳定)状态。究其原因, 主要是由于外部因素对浙江沿岸的底质破坏日益严重所致。
丁朋朋, 高春霞, 田思泉, 等, 2019. 浙江南部近海蟹类群落结构及其与环境因子的关系. 海洋渔业, 41(6): 652-662 |
马文刚, 夏景全, 魏一凡, 等, 2022. 三亚蜈支洲岛海洋牧场近岛区底表大型底栖动物群落结构及评价. 热带海洋学报, 41(3): 135-146 |
王金宝, 李新正, 王洪法, 等, 2018. 山东长岛潮间带大型底栖动物生态特征研究. 海洋科学, 42(10): 41-52 |
王航俊, 姚炜民, 林义, 等, 2020. 乐清湾大型底栖动物群落及其与环境因子之间的关系. 海洋学报, 42(2): 75-86 |
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. 海洋调查规范第6部分: 海洋生物调查: GB/T 12763.6-2007[S]. 北京: 中国标准出版社: 56-62.
|
卢占晖, 朱文斌, 徐开达, 等, 2018. 浙江沿岸海域春季螺类群落结构特征及其与环境因子的关系. 水生生物学报, 42(3): 606-615 |
刘迅, 王莉, 顾晓英, 等, 2017. 浙江檀头山岛周围海域夏、秋季大型底栖动物群落结构特征. 海洋与湖沼, 48(3): 553-567 |
刘瑞玉, 2008. 中国海洋生物名录. 北京: 科学出版社, 654-983
|
严润玄, 韩庆喜, 王晓波, 2020. 杭州湾和三门湾拖网大型底栖动物群落组成和多样性研究. 海洋与湖沼, 51(3): 484-493 |
李少文, 任中华, 王田田, 2017. 黄河口海域夏季大型底栖动物的生态学特征. 渔业科学进展, 38(6): 9-17 |
李亚芳, 杜飞雁, 谷阳光, 等, 2016. 雷州半岛东南部海域春季大型底栖动物群落特征及其与环境因素的关系. 南方水产科学, 12(6): 33-41 |
杨俊毅, 高爱根, 王永泓, 等, 2000. 台州列岛海域贝类种类组成与分布特征. 东海海洋, 18(1): 37-42 |
何贤保, 俞存根, 覃涛, 等, 2013. 舟山渔场及附近海域虾类群落结构特征分析. 水生生物学报, 37(3): 553-559 |
邹莉, 张龙, 张宾, 等, 2010. 浙江省沿岸岛礁区贝类种类组成与分布特征. 水产学报, 34(11): 1769-1775 |
陈万东, 伍尔魏, 蔡厚才, 等, 2017. 浙江玉环披山岩礁潮间带贝类种类数量组成与生态特征. 浙江海洋学院学报(自然科学版), 36(1): 1-8 |
陈伟峰, 彭欣, 叶深, 等, 2018. 浙南近海虾类群落结构及其多样性分析. 海洋科学, 42(3): 37-45 |
林建杰, 2022. 闽江口潮间带大型底栖动物群落基本特征分析. 渔业研究, 44(1): 33-43 |
金文育, 彭欣, 王宁, 等, 2017. 渔山列岛夏季潮间带大型底栖生物群落结构研究. 海洋科学, 41(3): 17-25 |
周细平, 李贞, 吴培芳, 等, 2019. 珠江河口区大型底栖动物群落结构. 生物多样性, 27(10): 1112-1121 |
赵凡淇, 刘卫霞, 隋吉星, 等, 2018. 秋季山东南部近岸海域大型底栖动物群落特征. 海洋科学进展, 36(1): 117-127 |
胡桂坤, 秦璐璐, 李郁郁, 等, 2019. 基于ABC曲线的天津潮间带生物群落受扰动的分析. 天津科技大学学报, 34(5): 57-62 |
祝超文, 张虎, 袁健美, 等, 2022. 南黄海潮间带大型底栖动物群落组成及时空变化. 上海海洋大学学报, 31(4): 950-960 |
袁伟, 王俊, 左涛, 等, 2020. 莱州湾大型底栖动物群落结构及其动态变化特征. 海洋学报, 42(6): 52-61 |
梁静香, 周永东, 王忠明, 等, 2020. 三门湾大型底栖动物群落结构及其与环境因子的关系. 应用生态学报, 31(9): 3187-3193 |
蔡如星, 黄惟灏, 刘月英, 等, 2001. 浙江动物志-软体动物. 杭州: 浙江科学技术出版社, 38-49
|
蔡林婷, 王一农, 李祥付, 等, 2013. 舟山东极岛潮间带贝类生态学初步研究. 海洋科学, 37(8): 47-54 |
AGLEN A, FLYN L, GODFLOR, et al, 1981. Surveys of the marine fish resources of Peninsular Malaysia, June-July 1980. Institute of Marine Research, Bergen, 9: 1-69 |
BRÖNMARK C, 1990. How do herbivorous freshwater snails affect macrophyte?-A comment. Ecology, 71(3): 1212-1215 |
CLARKE K R, WARWICK R M, 2001. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation. 2nd ed. Plymouth: PRIMER-E, 1-91
|
ELGER A, LEMOINE D, 2005. Determinants of macrophyte palatability to the pond snail Lymnaea stagnalis. Freshwater Biology, 50(1): 86-95 |
MARGALEF R, 1958. Information theory in ecology. General Systems, 3: 36-71 |
PINKAS L, OLIPHANT M S, IVERSON L K., 1971. Food habits of albacore, bluefin tuna, and bonito in California waters. Fish Bulletin, 152: 1-105 |
WARWICK R M, 1986. A new method for detecting pollution effects on marine macrobenthic communities. Marine Biology, 92(4): 557-562 |