 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 徐帆, 张晶, 赵亮. 2023.
- XU Fan, ZHANG Jing, ZHAO Liang. 2023.
- 基于分源物理生态耦合模型研究东海表层溶解无机氮对叶绿素季节变化的影响
- EFFECT OF DISSOLVED INORGANIC NITROGEN ON SEASONAL VARIATION OF SURFACE CHLOROPHYLL IN THE EAST CHINA SEA IN PHYSICAL-BIOLOGICAL COUPLING MODEL
- 海洋与湖沼, 54(2): 375-386
- Oceanologia et Limnologia Sinica, 54(2): 375-386.
- http://dx.doi.org/10.11693/hyhz20220500120
文章历史
-
收稿日期:2022-05-06
收修改稿日期:2022-06-06




2. 天津科技大学海洋资源化学与食品技术教育部重点实验室 天津 300457
2. Key Laboratory of Marine Resource Chemistry and Food Technology, Tianjin University of Science and Technology, Ministry of Education, Tianjin 300457, China
浮游植物是海洋初级生产的主要贡献者, 其生物量占全球总初级生产的近50% (Smith, 2006), 一方面它可以为异养生物提供能量来源(Chassot et al, 2010), 维持食物链的平衡; 另一方面通过生物泵作用促使海洋吸收大气二氧化碳, 调节气候变化(孙军等, 2016), 还与赤潮、低氧等环境问题紧密相关(宋洪军等, 2011)。叶绿素a (chl a)浓度是反映海洋浮游植物现存量和动态变化的重要指标(党晓岩等, 2019)。研究海洋中chl a的时空变化规律与影响机制对于认识浮游植物以及初级生产力分布特征, 评估生物资源蕴藏量及生产潜力、合理开发利用海洋生物资源和实行渔业生产农牧化、保护海洋环境安全等具有重要意义。
东海陆架连接着太平洋和东亚大陆, 其物理场和生态系统同时受到陆源河流和外源黑潮等多种因素的影响(Gong et al, 2003)1220, 具有复杂的水文和生态环境, 使东海成为典型的季节性层化陆架海生态系统(赵辉等, 2014)。汇入东海的河流携带着丰富的营养物质(Wang et al, 2015), 其中包括中国最大的河流——长江。剧烈的人类活动导致长江河口地区硝酸盐浓度在近年来增加了3倍, 赤潮等有害藻华在数量和规模上都急剧增加(Zhou et al, 2008), 且不同区域的浮游植物受到长江来源营养盐的影响存在较大差异性(许灵静, 2019)。大气干、湿沉降中氮输入占外源总输入的56%, 且氮沉降在中国东部海域支持100~200 mmol/(m2·a)的新初级生产力, 约为总新生产力的1.1%~3.9%, 是进入东海的一个重要的外部营养源(Chen et al, 1999; Zhang et al, 2010)。此外东海还接收来自台湾暖流和黑潮等环流带来的大洋信号的影响(张玉荣等, 2016)。黑潮来源营养盐对浮游植物的贡献在陆架中部、外陆架北部最显著且具有明显季节变化特征(王玉衡等, 2017)。台湾海峡流向东海输入的营养盐主要存在于夏季, 而冬季则相对较少(杨金湘等, 2018)。东海的多种营养盐来源使得东海生态系统的生产力非常高, 平均初级生产达到297~515 mg/(m2·d) (Gong et al, 2003)1232, 而且多种来源营养盐的共同作用也为东海浮游植物季节变化特征及空间分布带来了复杂的影响(Zhang et al, 2007; 文斐等, 2012; 张玉荣等, 2016; Hao et al, 2019)。
以往的研究已经对东海浮游植物的空间分布与季节变化等方面进行了系统分析, 然而受研究方法的局限往往无法区分不同来源溶解无机氮(dissolved inorganic nitrogen, DIN)的不同作用, 对认识东海浮游植物季节变化以及对DIN的响应机制带来了诸多困难。近年来, 在研究多来源与多机制作用的陆架海生态系统时, 一种对营养盐分源的生态动力学模型得到广泛应用。分源模型可以将DIN按照不同来源分别追踪, 完全显式表达某一来源DIN在生态系统中的物理输运和生化循环过程, 反映不同来源DIN对浮游植物在季节变化中的作用, 并直接得到它们各自所支持的浮游植物的真实变化。本文以添加分源模块的物理-生态耦合模型为主要手段, 分析东海中河流径流、大气沉降、台湾海峡流以及台湾以东的黑潮四个来源DIN所支持的表层chl a变化情况, 进而定量评估多源DIN对表层chl a季节变化的贡献, 并且解决各来源DIN在哪些海区起主导作用、作用途径是什么等问题。
1 数据与方法 1.1 分源物理生态耦合模型本文基于添加了分源模块的物理-生态耦合模型研究东海不同来源DIN对表层chl a季节变化的影响。其中物理-生态耦合模型用于计算总营养盐、浮游植物、碎屑等变量, 而营养盐分源示踪模块用于计算生态系统中来源不同的营养盐、浮游植物等在生态系统中的独立循环。
1.1.1 物理-生态耦合模型物理-生态耦合模型可以分为物理和生态两个模块。物理水动力模块基于普林斯顿海洋模型(princeton ocean model, POM) (Blumberg et al, 1987)建立, 空间分辨率为1/18°, 垂向分辨率为21层。模型区域覆盖了渤海、黄海和东海, 范围是24°~41°N, 117.5°~131.5°E, 本文主要研究为200 m等深线以浅的东海陆架区域, 即图 1中蓝色区域。此外物理模块还考虑了东海4个主要分潮的作用(M2、S2、K1和O1), 添加了10条河流径流, 本文研究区域包含4条(其中主要影响东海的为长江), 还引入了悬浮颗粒物以考虑其对光合作用的影响。
 |
| 图 1 物理-生态耦合模型中的东海陆架地形图 Fig. 1 The topography of the East China Sea shelf in physical-biological coupled model 注: 等值线分别为50, 100和200 m等深线; 黄色点表示河流入海位置(从北至南分别为淮河、长江、钱塘江、闽江); 红色直线代表南边界台湾海峡和黑潮的位置; 蓝色覆盖区域为本文研究区域 |
生态模块与Zhao等(2011)和Wang等(2019)中所使用的一致, 模型中物质流程以及主要生物参数可从中获取。其中包括三种营养盐: DIN、溶解无机磷(dissolved inorganic phosphorus, DIP)和硅酸盐(silicate, SIL); 两种浮游植物: 硅藻(diatom, DIA)和鞭毛藻(flagellate, FLA); 两种有机碎屑: 生物硅(biogentic silica, SIS)和生物硅之外的碎屑(detritus, DET)。主要的生物过程包括: 浮游植物作为海洋中的生产者, 利用太阳光进行光合作用制造有机物, 同时通过呼吸作用消耗有机物而还原营养盐; 营养盐作为光合作用的基础, 为浮游植物提供光合作用的原料。碎屑主要来自于生物死亡生成, 碎屑可以通过矿化作用补充海水中的营养盐。
1.1.2 营养盐分源模块分源营养盐模块与Zhang等(2019, 2021)中使用一致, 将生态系统中DIN这一整体依据其不同的来源分别进行独立计算, 同时考虑了其各自的物理过程和生态过程。东海的浮游植物在不同区域主要受到DIN或DIP的限制, 但DIP主要来自于黑潮, 而DIN的供给来源更加复杂多样且季节变化较强(Zhang et al, 2019), DIN对海洋环境的初级生产、生态系统功能和生物地球化学过程等也发挥着重要作用(Worsfold et al, 2008; Yu et al, 2012; 蒋昊等, 2020), 故本文将对东海DIN进行分源。分源模块中与DIN有关的生态变量, 即DIN、DIA、FLA和DET, 均按照其不同来源标记。分源模块中每增加一个外源种类, 就将增加一套完整的DIN、DIA、FLA、DET的控制方程来独立求解这一来源的生态变量。这一做法可以反映实际海洋生态系统中的分源情况, 但也会使计算时间成倍增加。
模型中的4种外源DIN被标记并追踪, 它们分别为: 河流径流(rivers, R), 包括淮河、长江、钱塘江和闽江; 大气沉降(atmospheric deposition, A), 包括大气干沉降和湿沉降; 台湾海峡(Taiwan Strait, T)和台湾以东黑潮(Kuroshio, K)。其中河流入海口浓度数据来自Zhang(1996)和Liu等(2009), 大气干湿沉降气候态分布数据来自Zhang等(2011)。台湾海峡来源的营养盐、浮游植物和碎屑数据来自陈镇东教授(个人通讯)。黑潮来源的营养盐、浮游植物和碎屑数据来自日本气象厅(Japan Meteorological Agency)多年长期观测的平均值。该模拟反映了2000年以前东海低营养生态系统的气候态条件。海底水-沉积物界面过程对于各个来源的生态变量来说都是独立的, 比如有机物的沉降和沉积物的矿化等, 囊括在这一来源的物质循环中, 从而底层不作为一个新的营养盐外源。下文中使用DINR、DINA、DINT、DINK来分别代指来自这4种来源的营养盐, 总营养盐被记做DINW。不同来源DIN支持的chl a (下文简称为不同来源的chl a)也采用类似的标记方法, 分别表示为chlR、chlA、chlT、chlK以及总chl a浓度chlW。
分源模块中生物过程引起的源汇项依据各分源DIN浓度占总DIN浓度的比例来分配。主要生态变量DIN, DIA, FLA, DET的控制方程如下:
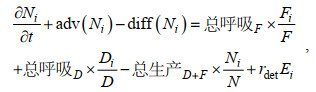 (1)
(1)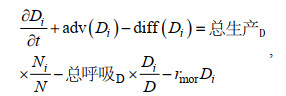 (2)
(2) (3)
(3)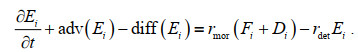 (4)
(4)其中, N代表溶解无机氮(DIN), D为硅藻(DIA), F为鞭毛藻(FLA), E为碎屑(DET), i代表某一种DIN来源, adv和diff代表变量的对流项和扩散项, rmor为浮游植物死亡系数, rdet为碎屑分解系数, 均与温度有关。该方法保证分源前后的物质守恒性, 即分源后的变量之和与不分源时的总变量相等。本文将分源模块单独应用于DIN算例, 将4种外部来源营养源对应的DIN浓度、浮游植物和碎屑的初始条件设为0, 并在气候态条件下运行了5 a直到状态变量达到稳定状态, 并使用最终年份的结果进行分析。
1.2 模式验证水动力模块在Guo等(2006)以及Wang等(2008)中进行了详细验证。对于生态模块, Zhao等(2011)对比模型在PN断面的营养盐和chl a结果与观测吻合, 春季, 随着浮游植物的生长, 表层的营养盐迅速消耗殆尽; 夏季表层营养盐浓度降低, 限制了浮游植物的生长; 秋季, 由于表底混合加剧, 表层营养盐浓度略有升高, 浮游植物在中层仍有较高的浓度。Wang等(2019)将chl a模拟结果与卫星结果对比验证发现, 在春季和夏季东海内陆架的表层chl a浓度较高, 而秋季和冬季则较低, 并且外陆架表层chl a的低值在模型与遥感结果中也均被证实。另外Zhang等(2019)将模型chl a和DIN结果与观测结果的垂直分布对比验证显示, 冬季, chl a浓度在外陆架上层的较高, 内陆架和中陆架相对较低。而DIN浓度在垂直方向上较为均匀; 夏季, 最大chl a深度逐渐向东下降, DIN浓度表现为向海岸倾斜的垂直变化。结果均与观测相似。这充分说明了该模型可以成功再现东海chl a的空间分布和季节变化。
为进一步验证模型数据在东海表层chl a季节变化方面的可靠性, 本文将东海空间平均chl a浓度季节变化和常见的卫星chl a产品(表 1)进行对比验证(图 2)。chl a的浓度在1月时模式结果与各卫星数据基本都为最低, 随后快速增加, 到春季4月分源模式结果与各卫星数据都达到最高, 随后又开始快速下降直至夏、秋季, chl a浓度变化趋于稳定不再剧烈升降。从总体季节变化趋势来看, 模型模拟的东海表面chl a浓度与前人遥感方法得出的结果均在4月出现最高值, 1月出现最低值, 表明模式与卫星结果的chl a浓度季节变化趋势基本吻合。从数值范围来看, 不同数据的结果之间均存在一定的偏差, 模型结果大致处于前人研究结果数据范围内, 仅在冬季时存在一定数值上差异, 这可能是因为模型中表层chl a仅采用200 m以浅区域的平均, 与前人文献中的范围不完全相同所致。
| 数据类别 | 数据详情 | 数据来源 |
| SeaWiFS数据 | 1997年10月至2007年5月SeaWiFS卫星三级反演数据 | 伍玉梅等(2008)中fig. 1 |
MODIS重构数据 |
2003年1月至2012年12月, 空间分辨率是2.5′ | 郭俊如(2014)中fig. 4~8 |
| 标准叶绿素数据与LMC数据 | 标准叶绿素数据来源于OC4算法计算的OC-CCI叶绿素产品, LMC为基于局部修正的叶绿素产品 | Hao等(2019)附录fig. 2 |
| 注: SeaWiFS为海视宽视野传感器, MODIS为中分辨率成像光谱仪, LMC为基于水反射率和标准叶绿素数据误差之间的经验关系计算所得局部修正的叶绿素a数据集, OC-CCI为海洋水色气候变化倡议, OC4为经验海洋颜色叶绿素算法第4版 | ||
 |
| 图 2 模型与卫星chl a浓度在东海表层的季节变化 Fig. 2 Seasonal variation in chlorophyll concentration in the East China Sea surface from model and satellite data 注: 图中黑色虚线为分源模型结果, 误差棒为月内变化标准差 |
东海陆架表层DIN浓度水平梯度大, 各来源DIN影响的海域范围与程度有所差异, 故探明它们各自支持的chl a在东海表层的时空分布, 对于了解不同来源DIN对chl a季节变化的影响具有重要意义。不同来源chl a各季节空间分布如图 3所示, 其中2月、5月、8月、11月分别代表冬季、春季、夏季、秋季。
 |
| 图 3 东海4种来源DIN支持的表层chl a浓度以及chl a总浓度在2月, 5月, 8月, 11月的空间分布 Fig. 3 The spatial distributions of surface chlorophyll concentration from four sources of DIN and their sum over the East China Sea in February, May, August, and November 注: chlR为河流来源DIN所支持的chl a, chlK为黑潮来源的DIN所支持的chl a, chlT为台湾海峡来源的DIN所支持的chl a, chlA为大气来源的DIN所支持的chl a, chlW为总chl a, 空白区域表示非研究区域的其他海域 |
chlR主要聚集在内陆架附近, 在春、夏季的长江口及其以北的海域浓度最大, 几乎覆盖50 m等深线以浅的广阔区域; 冬季、秋季时, 长江口附近区域依旧存在高值, 但高值区域从秋季开始逐渐缩减, 到冬季时最弱, 存在沿岸边狭窄区域向南延伸的趋势, 一路到达台湾海峡。chlA春季相对整体浓度最高并且分布最广泛, 几乎覆盖整个东海陆架区域; 冬季浓度最低, 分布范围最小, 仅有小部分出现在朝鲜半岛南部及沿岸区域; 夏季主要分布在内陆架且在南部存在由近岸向外海延伸趋势; 到秋季近岸区浓度减弱, 主要分布在外陆架及北部地区。chlK在冬季浓度最高且分布集中于东海外陆架海区; 春季开始向内陆架及沿岸区域延伸, 扩散至台湾岛北部以及长江口及其北部区域, 并且几乎覆盖整个陆架; 夏季分布范围最小, 仅存在于台湾东北及长江口附近; 秋季又逐渐回归外陆架区域。chlT在冬季浓度极低, 仅出现在朝鲜半岛南部小部分区域, 春、夏季主要分布在台湾海峡并向东北延伸, 秋季延伸到了外陆架。
对于chlW, 冬季主要分布于沿岸以及外陆架区域, 这主要来自于chlR和chlK的贡献, 分别归因于长江冲淡水(Changjiang diluted water, CDW)在台湾暖流内侧向南流动到达台湾海峡(Chang et al, 2003)15-3和黑潮在台湾东北的表层入侵分支较强(Hsin et al, 2013)。随着温度的上升, chlW在春季浓度达到最高, 内、外陆架都达到较高水平, chlR为主要贡献者, 其他三种来源chl a也开始增加。因为春季河流持续向东海输送营养盐, 尤其是河流来源的主力军长江, CDW在春季时离岸流动, 能到达黑潮锋区(Chang et al, 2003)15-3, 这为东海带来了大量的陆源营养盐以支持初级生产, 但是chlW在中陆架北部则相对较低, 这是因为该区域4种来源的影响均较弱, 再加上这可能与早春水华对表层营养盐的消耗也有关(Zhang at al, 2019); 夏季chlW在内陆架以及中陆架南部的高浓度主要来自chlR以及chlA的贡献, 这与夏季层化加强导致底层积累的营养盐无法对表层海水中消耗的营养盐进行补充有关(王文涛, 2013), 而且夏季较强的CDW自东海北部向东北方向移动(Lie et al, 2003; Hu et al, 2016), 因此陆源输入是夏季海水营养盐的主要来源。另外台湾海峡流在夏季向东海的持续运输(杨金湘等, 2018), 以及黑潮的近岸分支在夏季可以从陆坡处缓慢涌升至长江口外区(Yang et al, 2012), 导致chlT和chlK也有不弱的贡献; 秋季时chlW高值区域向沿岸区域与外陆架转移, 其中沿岸区主要因为秋季东北季风形成, CDW撤回到沿岸区域(Chang et al, 2003), chlR也因此向岸缩减, 而外陆架主要由于chlK显著增加。
为进一步明确不同来源DIN对东海表层chl a季节变化的影响, 本文计算得出各来源DIN所支持chl a浓度季节变化如图 4所示。首先chlW, 冬季1月浓度最低为0.29 mg/m3, 此时的4种来源中除chlK外都处于全年的最低水平。随着4种chl a浓度在1~4月的快速增长, chlW也开始快速上升到春季4月升至最高值1.21 mg/m3, 春季4月后chlK开始急剧下降, 导致chlW开始下降, 5月后chlR、chlT以及chlA也开始下降, 导致chlW加速下降。6月后主要因为chlT开始上升, chlW的下降放缓。直至8月形成一个小低谷后, chlK和chlA开始迅速增长, 导致chlW有所抬升。到秋季10月形成另一个小高峰后, 除chlK的其余三种chl a浓度都呈现下降趋势, 导致chlW也开始下降。总体来看, chlW呈现双峰式分布, 且从高到低分布依次为: 春季, 秋季, 夏季, 冬季。
 |
| 图 4 东海表层各来源DIN所支持的叶绿素a浓度季节变化 Fig. 4 Seasonal variations of surface chl a concentration supported by DIN from four sources in the East China Sea |
对于4种来源的chl a浓度, chlK与chlR先后占据主导地位。从冬季12月起至春季4月, 一直都是chlK主导并且呈现快速上升的趋势, 从5月开始到11月chlR转为主导。chlR在冬季1月最低仅0.057 mg/m3, 随后开始增长, 直至春季5月升至最高0.42 mg/m3, 而后在夏季以及秋季都表现出下降的整体趋势。且chlR从高到低分布依次为: 春季, 夏季, 秋季, 冬季。其次chlA也占有重要地位, 同样在冬季1月达到最低值0.038 mg/m3, 春季5月上升至最高值0.29 mg/m3左右。夏季开始下降, 8月形成小低谷后在秋季开始攀升, 总体来说, chlA季节变化呈现双峰式分布, 且从高到低分布依次为: 春季, 秋季, 夏季, 冬季。对于chlK来说, 冬季浓度相对其他来源最高, 且呈现上升趋势。春季继续加速上升到4月达到最高为0.42 mg/m3左右。随后下降至夏季8月达到最低值为0.051 mg/m3左右。秋季开始逐步上升。从高到低分布依次为: 春季, 冬季, 秋季, 夏季。chlT整体浓度最低, 冬季1月最低为0.013 mg/m3。在春季5月达到最高值0.13 mg/m3, 在夏季8月又迎来一个小高峰之后秋季开始逐渐降低。chlT季节变化也有双峰式分布特点, 且从高到低分布依次为: 春季, 夏季, 秋季, 冬季。综上所述, 除chlR外其余3种来源chl a均呈双峰结构, 且春季为各来源chl a的增长爆发期, chlK在4月达到最高峰值, 而其他3种来源也都在5月达到了最高峰值。chlW在1月达到最低也与chlR、chlA、chlT在1月时最低密切相关。
2.2 不同来源chl a主导区域影响研究区域的4条主要河流与大气干湿沉降携带着陆源信号从西部进入东海, 而由沿岸流、南海水以及黑潮分支构成的台湾海峡水(Hu et al, 2010)与黑潮主轴则携带大洋信号由南部进入东海。4种不同来源的DIN供给东海浮游植物生长的时间与空间差异, 导致不同来源chl a的主要影响区域也各不相同。本文将每个季节3个月chl a浓度平均, 并将不同来源的季节平均chl a进行了比较, 定义某一来源chl a在4种来源中最高浓度所占区域其主导区域, 不同来源chl a的主导区域如图 5所示。
 |
| 图 5 东海不同来源chl a主导区域在冬季, 春季, 夏季, 秋季的分布 Fig. 5 The distributions of chl a-dominated areas from different sources in the East China Sea in winter, spring, summer, and autumn |
冬季, chlR主导区域占据长江口及其北部的内陆架区域, 且闽浙沿岸流冬季沿岸向西南流动(Qiao et al, 2006), 使其一直延伸至台湾海峡西侧的狭长沿岸区域; chlA所主导的区域主要占据济州岛以西的狭小海域; chlK主导区域在冬季最大, 占据整个外陆架区域, 并且向岸入侵也达到最强, 且在28°~31°N之间的入侵有达到长江口外部区域的趋势, 其北部的向岸入侵也达到了对马海峡区域; chlT主导区域在冬季最小, 主要占据台湾西北小片海域, 这可能与台湾海峡流在冬季强季风的影响相关(Zhang et al, 2009)。春季, chlR主导区域占据整个沿岸地区, 初具向东延伸的趋势, 这与长江等河流的在春季时离岸流动有关(Chang et al, 2003)15-8。chlA主导区域主要占据在中陆架北部并向南延伸, 属于河流径流以及黑潮影响较弱的区域。chlK主导外陆架狭长海区, 由台湾北部一直延伸至对马海峡, 相比冬季有向外陆架退去的趋势。chlT主导区域在春季的向东北延伸至东海中部。夏季, chlR主导区域在长江口外继续离岸延伸至济州岛西, 但由于夏季沿岸流转向东北(Lü et al, 2006), 南部沿岸区域开始缩减。chlA主导区域相比春季向外陆架延伸, 且所占范围增至最大。chlK所主导的区域在夏季最小, 主要占据在台湾东北部的小块区域, 这可能与黑潮表层向岸输运在夏季最小有关(Oey et al, 2010; Wu et al, 2014)。chlT夏季的主导区域达到最大, 从台湾海峡向东北延伸至中陆架海区。秋季, chlR主导区域开始向岸回撤, 东海南部沿岸又逐渐恢复主导区域, 这与秋季的东北季风的形成和沿岸流的影响有关(Chang et al, 2003)。chlK主导的区域在外陆架又开始扩张。chlT所主导的区域相比夏季缩减, 主要被黑潮主导区域的扩张所侵占。
本文定义的主导区域, 能够直观地表现出不同来源chl a在空间上集中分布的变化情况, 但是局限于计算方法, 主导区域内主导来源因素能否占据绝对贡献地位以及其他非主导来源能贡献多少尚未可知, 所以定量评估不同主导区域内的各来源贡献情况是有必要的。表 2分别为4个季节不同主导区域中的各来源chl a浓度占比。chlR主导区域主要分布在内陆架, 作为主导因素chlR贡献在各季节占比均超过60%, 处于较强的主导地位, 除主导因素外, 还受到chlA的较强影响, 占比在18%~29%之间; chlK主导区域集中在外陆架, 在冬季主导因素chlK呈垄断地位, 占比高达82%。春、夏、秋则受到chlA和chlT的一定影响, 占比分别在15%~23%之间和7%~16%。chlR远离外陆架所以几乎不参与贡献; chlT主导区域主要在台湾海峡及中陆架, 冬季主导因素chlT贡献最高达62%, 而在秋季主导地位最弱为49%, 其主导区域主要还受到chlA的贡献, 各季节占比在15%~29%之间, 以及冬、春、秋季也受到chlK较强贡献, 占比达到13%~20%; chlA主导区域主要在东海北部, 但chlA的主导地位也相对较弱, 占比在46%~54%之间, 并且在四季主导区域内均容易受到其他3个来源的影响。我们发现各主导区域内除了主导因素, 还受到其他来源不同程度的影响。另外在冬、春、夏、秋季4种来源主导区域内主导因素的贡献占比均达到了46%以上, 而且各主导区域内的非主导因素均在29%以下, 也进一步说明各来源主导分区是合理的。
| 季节 | 主导区域 | 不同源chl a浓度 | |||
| chlR | chlK | chlT | chlA | ||
| 冬季 | chlR主导区域 | 70% | 4% | 4% | 22% |
| chlK主导区域 | 2% | 82% | 6% | 10% | |
| chlT主导区域 | 8% | 16% | 62% | 15% | |
| chlA主导区域 | 22% | 16% | 10% | 52% | |
| 春季 | chlR主导区域 | 65% | 5% | 6% | 24% |
| chlK主导区域 | < 1% | 77% | 7% | 15% | |
| chlT主导区域 | 11% | 13% | 55% | 21% | |
| chlA主导区域 | 19% | 17% | 15% | 49% | |
| 夏季 | chlR主导区域 | 71% | 5% | 6% | 18% |
| chlK主导区域 | 3% | 65% | 9% | 23% | |
| chlT主导区域 | 11% | 4% | 58% | 28% | |
| chlA主导区域 | 10% | 16% | 20% | 54% | |
| 秋季 | chlR主导区域 | 69% | 7% | 6% | 29% |
| chlK主导区域 | 2% | 64% | 16% | 18% | |
| chlT主导区域 | 2% | 20% | 49% | 29% | |
| chlA主导区域 | 18% | 20% | 16% | 46% | |
| 注: chlR为河流来源DIN所支持的chl a; chlK为黑潮来源的DIN所支持的chl a; chlT为台湾海峡来源的DIN所支持的chl a; chlA为大气来源的DIN所支持的chl a | |||||
不同来源chl a的季节变化十分复杂, 其生长受到光照、温度和营养盐的共同控制(Zhang et al, 2019)。但是对于表层浮游植物来说, 光照并不是其生长的限制因素。而对于温度来说, 不同chl a来源主导区域内温度季节变化十分相似(图 6), 这说明温度也并非造成不同来源chl a季节变化差异的决定性因素, 但是在同一时间不同区域的温度有一定差异也对各来源chl a形成了一定影响, 例如在冬季、春季时黑潮主导区域与台湾海峡主导区域内的温度相对较高, 为浮游植物的生长提供了更加适宜的环境, 而此时的河流以及大气沉降来源主导区域内温度较低, 就会对浮游植物的生长造成影响。到了夏季和秋季, 各主导区域内温度差异较小, 此时温度对浮游植物生长的限制也达到最小。营养盐由开边界与底边界供给、环流、混合和分层(Wang et al, 2015)等物理过程控制, 又受到水体内的生物过程调制(Chen et al, 2021), 从而在时空分布上存在强烈的季节变化, 这在很大程度上造成了不同来源chl a的季节变化差异。
 |
| 图 6 东海表层不同来源chl a主导区域内温度季节变化 Fig. 6 Seasonal variations of temperature in chl a-dominated areas from different sources in the surface East China Sea 注: tempR为河流来源chl a主导区域内的平均温度; tempK为黑潮来源chl a主导区域内的平均温度; tempT为台湾海峡来源chl a主导区域内的平均温度; tempA为大气来源chl a主导区域内的平均温度; tempW为东海平均温度 |
为了进一步在季节尺度上明确DIN对chl a的影响, 本文计算得出了东海表层总DIN以及河流径流、黑潮、台湾海峡、大气沉降4种不同来源的DIN季节变化(图 7), 并与它们各自支持的chl a季节变化进行对比。对于DINW来说, 冬季表层DINW浓度很高, 但chlW却最低, 说明此时浮游植物增长受到温度限制, 在2月DINW达到了最高值7.69 mmol/m3, 春季3月chlW开始快速增长, 导致DINW下降。到5月DINW形成小低谷浓度为5.49 mmol/m3, 对应的chlW也增至高值; 夏季小幅度增长到7月形成一个小高峰浓度达到6.23 mmol/m3, 此时对应chlW的缓慢下降; 随后DINW又快速下降, 而chlW缓慢上升, 到秋季9月DINW降到全年最低水平为4.63 mmol/m3, 而chlW又即将迎来一个小高峰。而后DINW快速上升, chlW又开始下降。综上我们发现, DINW和chlW均呈现双峰型季节变化, 二者高峰与低谷相互对应, 说明chlW的增长与DINW的消耗之间存在很好的响应关系。
 |
| 图 7 东海表层各来源DIN季节变化 Fig. 7 Seasonal variations of surface DIN from four sources in the East China Sea 注: DINR为河流来源DIN, DINK为黑潮来源的DIN, DINT为台湾海峡来源的DIN, DINA为大气来源的DIN, DINW为总DIN |
对于4种不同来源的DIN来说, DINR在4种外部来源中全年占据主导地位, 在冬季呈缓慢上升的趋势, 初春3月DINR达到最高值3.44 mmol/m3, 此时chlR也进入快速增加的阶段, 并且到5月达到了最高。随后DINR下降到5月形成小低谷, 夏季浓度回升, 7月达到高峰后继续降低, 到9月降至最低值2.56 mmol/m3, 而chlR短暂上升后也迎来了下降, DINR的主导地位也使得chlR在多半时间占据主导地位。DINA浓度在4种来源中也相对较高, 冬季缓慢上升到春季3月, 对应chlA也开始增长。后DINA缓慢下降至5月, 夏季6月增至最高2.09 mmol/m3, 随后缓慢下降至秋季10月达到最低1.25 mmol/m3, 此期间使得chlA也成为除chlR外最重要的来源。DINK冬季1月最高达1.56 mmol/m3, 后开始被浮游植物利用而快速下降, chlK快速增长。后因海水层化影响, 表层DINK减少, 到6月降至最低0.29 mmol/m3左右, 一直到8月持续保持较低浓度, 同样chlK也随之降低。DINT全年保持较低水平, 且季节变化相对不明显, 其浓度保持在0.35~0.63 mmol/m3之间, chlT同样全年保持相对较低的水平。综上, DIN的季节变化在很大程度上影响着chl a的季节变化, 且不同来源之间的响应时间与程度有较大差异。
3 结论本文使用物理-生态耦合模型和分源模块, 分析了东海陆架区域中4个主要来源表层DIN所支持的表层chl a的季节变化, 并从不同角度讨论了不同来源DIN对东海表层chl a季节变化的影响。
东海表层chlK与chlR先后占据主导地位。春季时各来源chl a的均快速增长, 4种来源chl a在4~5月先后增至峰值, 除chlK在8月降至最低外其他三种来源chl a均在冬季1月达最低值。而且除了chlR外, 其余3种来源chl a均呈双峰结构。
不同来源的DIN所支持的chl a在不同季节上存在空间分布差异。chlR主要聚集在内陆架附近, 春、夏季分布较广, 而冬季和秋季逐渐缩减。chlA冬季最弱, 春季最强, 几乎覆盖整个东海陆架区域, 夏季主要分布在内陆架及东海南部, 秋季有向外陆架转移的趋势。chlK冬、秋季分布集中于外陆架海区, 春季向内陆架延伸, 夏季分布较少。chlT在冬季分布极少, 在春、夏季主要分布在台湾海峡并向东北延伸, 秋季延伸至外陆架。整体来看, 冬季CDW在台湾暖流内侧向南流动以及黑潮在台湾东北的表层入侵分支, 使得chlR和chlK较强, 造成chlW分布于沿岸以及外陆架区域。春季随着温度的上升, 且海水分层尚未完全形成, 各来源DIN能较有效的对chl a形成供给, 使得chlW在内、外陆架都达到较高水平。夏季表、底层海水之间的温度和密度差异导致底层DIN难以对表层进行补充, 且台湾海峡流东海的持续运输, 以及黑潮的近岸分支向岸涌升, 使得chlW内陆架以及中陆架南部较高。秋季东北季风形成, CDW撤回到沿岸区域, 再加上黑潮向东移动, 使得chlW分布向沿岸区域与外陆架转移。
各来源chl a主导区域大小和范围也呈现季节变化特征。冬季chlK主导区域最大, 其他3种来源达到最小。春季chlR主导区域向离岸延伸, chlT和chlA主导区域向中陆架延伸, chlK主导区域急剧向外陆架收缩。夏季chlR主导区域达到最大, 离岸延伸最远, 而chlK来源主导区域缩至最小, chlT也达到最大, chlA主导区域逐渐被其他来源挤占。秋季chlR、chlA以及chlT主导区域都开始收缩, chlK主导区域又开始向陆延伸。
另外, 不同来源chl a的主导区域内除了主导来源, 还受到其他来源chl a不同程度的贡献, 这主要与不同源chl a的空间分布有关。chlK在其主导区域内较强势, 其他来源chl a影响较少; chlR主导区内还存在chlA的一定贡献; chlT主导区域内在春、夏季主要还受到chlA的贡献, 秋季chlK贡献也有所增加, chlA主导区域内在四季均受到其他三个来源chl a的影响。
不同来源表层chl a季节变化差异主要受到DIN的影响, 且不同来源的chl a与DIN之间的响应时间与程度有较大差异。同时温度在一定程度上会影响chl a的生长和DIN对浮游植物的供给。
王文涛, 2013. 中国东海、黄海和渤海微表层营养盐分布及富集研究[D]. 青岛: 中国海洋大学: 38-42.
|
王玉衡, 赵亮, 沈家葳, 2017. 黑潮营养盐输入对东海陆架浮游生态系统影响的模型研究. 海洋与湖沼, 48(6): 1346-1354 |
文斐, 孙晓霞, 郑珊, 等, 2012. 2011年春、夏季黄、东海叶绿素a和初级生产力的时空变化特征. 海洋与湖沼, 43(3): 438-444 |
伍玉梅, 徐兆礼, 崔雪森, 等, 2008. 1997—2007年东海叶绿素a质量浓度的时空变化分析. 环境科学研究, 21(6): 137-142 |
许灵静, 2019. 长江口及其邻近海域黑潮次表层水的入侵机制及其生态效应[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 74-80.
|
孙军, 李晓倩, 陈建芳, 等, 2016. 海洋生物泵研究进展. 海洋学报, 38(4): 1-21 DOI:10.3969/j.issn.0253-4193.2016.04.001 |
杨金湘, 王佳, 2018. 台湾海峡冬、夏季氮通量的数值模拟研究. 海洋学报, 40(4): 30-40 DOI:10.3969/j.issn.0253-4193.2018.04.003 |
宋洪军, 季如宝, 王宗灵, 2011. 近海浮游植物水华动力学和生物气候学研究综述. 地球科学进展, 26(3): 257-265 |
张玉荣, 丁跃平, 李铁军, 等, 2016. 东海区叶绿素a和初级生产力季节变化特征. 海洋与湖沼, 47(1): 261-268 |
赵辉, 张淑平, 2014. 中国近海浮游植物叶绿素、初级生产力时空变化及其影响机制研究进展. 广东海洋大学学报, 34(1): 98-104 |
党晓岩, 伍玉梅, 樊伟, 等, 2019. 黄、东海Chl a浓度与环境因子的相关分析. 中国海洋大学学报, 49(S1): 93-101 |
郭俊如, 2014. 东中国海遥感叶绿素数据重构方法及其多尺度变化机制研究[D]. 青岛: 中国海洋大学: 69-70.
|
蒋昊, 赵亮, 张晶, 2020. 黄海溶解无机氮及颗粒有机氮收支与转化模型. 中国环境科学, 40(9): 3981-3991 |
BLUMBERG A F, MELLOR G L, 1987. A description of a three-dimensional coastal ocean circulation model [M] // HEAPS N S. Three-Dimensional Coastal Ocean Models, Volume 4. Washington: American Geophysical Union: 1-16.
|
CHANG P H, ISOBE A, 2003. A numerical study on the Changjiang diluted water in the Yellow and East China Seas. Journal of Geophysical Research: Oceans, 108(C9): 3299 |
CHASSOT E, BONHOMMEAU S, DULVY N K, et al, 2010. Global marine primary production constrains fisheries catches. Ecology Letters, 13(4): 495-505 |
CHEN D X, LIU Q Q, YIN K D, 2021. Numerical study of the three gorges dam influences on chlorophyll-a in the Changjiang estuary and the adjacent East China Sea. Journal of Ocean University of China, 20(1): 10-22 |
CHEN C T A, WANG S L, 1999. Carbon, alkalinity and nutrient budgets on the East China Sea continental shelf. Journal of Geophysical Research: Oceans, 104(C9): 20675-20686 |
GONG G C, WEN Y H, WANG B W, et al, 2003. Seasonal variation of chlorophyll a concentration, primary production and environmental conditions in the subtropical East China Sea. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 50(6/7): 1219-1236 |
GUO X Y, MIYAZAWA Y, YAMAGATA T, 2006. The Kuroshio onshore intrusion along the shelf break of the East China Sea: the origin of the Tsushima warm current. Journal of Physical Oceanography, 36(12): 2205-2231 |
HAO Q, CHAI F, XIU P, et al, 2019. Spatial and temporal variation in chlorophyll a concentration in the Eastern China Seas based on a locally modified satellite dataset. Estuarine, Coastal And Shelf Science, 220: 220-231 |
HSIN Y C, QIU B, CHIANG T L, et al, 2013. Seasonal to interannual variations in the intensity and central position of the surface Kuroshio east of Taiwan. Journal of Geophysical Research: Oceans, 118(9): 4305-4316 |
HU J Y, KAWAMURA H, L I C Y, et al, 2010. Review on current and seawater volume transport through the Taiwan Strait. Journal of Oceanography, 66(5): 591-610 |
HU Z F, WANG D P, PAN D L, et al, 2016. Mapping surface tidal currents and Changjiang plume in the East China Sea from geostationary ocean color imager. Journal of Geophysical Research: Oceans, 121(3): 1563-1572 |
LIE H J, CHO C H, LEE J H, et al, 2003. Structure and eastward extension of the Changjiang River plume in the East China Sea. Journal of Geophysical Research: Oceans, 108(C3): 3077 |
LIU S M, HONG G H, ZHANG J, et al, 2009. Nutrient budgets for large Chinese estuaries. Biogeosciences, 6(10): 2245-2263 |
LÜ X G, QIAO F L, XIA C S, et al, 2006. Upwelling off Yangtze River estuary in summer. Journal of Geophysical Research: Oceans, 111(C11): C11S08 |
OEY L Y, HSIN Y C, WU C R, 2010. Why does the Kuroshio northeast of Taiwan shift shelfward in winter?. Ocean Dynamics, 60(2): 413-426 |
QIAO F L, YANG Y Z, LÜ X G, et al, 2006. Coastal upwelling in the East China Sea in winter. Journal of Geophysical Research: Oceans, 111(C11): C11S06 |
SMITH V H, 2006. Responses of estuarine and coastal marine phytoplankton to nitrogen and phosphorus enrichment. Limnology and Oceanography, 51(1part2): 377-384 |
WANG Q, GUO X Y, TAKEOKA H, 2008. Seasonal variations of the Yellow River plume in the Bohai Sea: a model study. Journal of Geophysical Research: Oceans, 113(C8): C08046 |
WANG Y C, GUO X Y, ZHAO L, et al, 2019. Seasonal variations in nutrients and biogenic particles in the upper and lower layers of East China Sea Shelf and their export to adjacent seas. Progress in Oceanography, 176: 102138 |
WANG Y, JIANG H, JIN J X, et al, 2015. Spatial-temporal variations of chlorophyll-a in the adjacent sea area of the Yangtze River estuary influenced by Yangtze River discharge. International Journal of Environmental Research and Public Health, 12(5): 5420-5438 |
WORSFOLD P J, MONBET P, TAPPIN A D, et al, 2008. Characterisation and quantification of organic phosphorus and organic nitrogen components in aquatic systems: a review. Analytica Chimica Acta, 624(1): 37-58 |
WU C R, HSIN Y C, CHIANG T L, et al, 2014. Seasonal and interannual changes of the Kuroshio intrusion onto the East China Sea Shelf. Journal of Geophysical Research: Oceans, 119(8): 5039-5051 |
YANG D Z, YIN B S, LIU Z L, et al, 2012. Numerical study on the pattern and origins of Kuroshio branches in the bottom water of southern East China Sea in summer. Journal of Geophysical Research: Oceans, 117(C2): C02014 |
YU Y, SONG J M, LI X G, et al, 2012. Distribution, sources and budgets of particulate phosphorus and nitrogen in the East China Sea. Continental Shelf Research, 43: 142-155 |
ZHANG J, 1996. Nutrient elements in large Chinese estuaries. Continental Shelf Research, 16(8): 1023-1045 |
ZHANG J, GUO X Y, ZHAO L, 2019. Tracing external sources of nutrients in the East China Sea and evaluating their contributions to primary production. Progress in Oceanography, 176: 102122 |
ZHANG J, GUO X Y, ZHAO L, 2021. Budget of riverine nitrogen over the East China Sea shelf. Environmental Pollution, 289: 117915 |
ZHANG W Z, HONG H S, SHANG S P, et al, 2009. Strong southward transport events due to typhoons in the Taiwan Strait. Journal of Geophysical Research: Oceans, 114(C11): C11013 |
ZHANG J, LIU S M, REN J L, et al, 2007. Nutrient gradients from the eutrophic Changjiang (Yangtze River) Estuary to the oligotrophic Kuroshio waters and re-evaluation of budgets for the East China Sea Shelf. Progress in Oceanography, 74(4): 449-478 |
ZHANG Y, YU Q, MA W C, et al, 2010. Atmospheric deposition of inorganic nitrogen to the eastern China seas and its implications to marine biogeochemistry. Journal of Geophysical Research: Atmospheres, 115(D7): D00K10 |
ZHANG J, ZHANG G S, BI Y F, et al, 2011. Nitrogen species in rainwater and aerosols of the Yellow and East China seas: effects of the east Asian monsoon and anthropogenic emissions and relevance for the NW Pacific Ocean. Global Biogeochemical Cycles, 25(3): GB3020 |
ZHAO L, GUO X, 2011. Influence of cross-shelf water transport on nutrients and phytoplankton in the East China Sea: a model study. Ocean Science, 7(1): 27-43 |
ZHOU M J, SHEN Z L, YU R C, 2008. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River. Continental Shelf Research, 28(12): 1483-1489 |