 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 陆化杰, 何静茹, 陈静, 崔红, 欧玉哲, 赵懋林, 陈新军. 2023.
- LU Hua-Jie, HE Jing-Ru, CHEN Jing, CUI Hong, OU Yu-Zhe, ZHAO Mao-Lin, CHEN Xin-Jun. 2023.
- 不同气候年间西北印度洋鸢乌贼角质颚微结构及生长特性差异
- DIFFERENCES IN BEAK MICROSTRUCTURE AND GROWTH CHARACTERISTICS OF STHENOTEUTHIS OUALANIENSIS IN THE NORTHWEST INDIAN OCEAN BETWEEN DIFFERENT YEARS
- 海洋与湖沼, 54(2): 514-525
- Oceanologia et Limnologia Sinica, 54(2): 514-525.
- http://dx.doi.org/10.11693/hyhz20220700185
文章历史
-
收稿日期:2022-07-12
收修改稿日期:2022-09-20



2. 自然资源部海洋生态监测与修复技术重点实验室 上海 201306;
3. 国家远洋渔业工程技术研究中心 上海海洋大学 上海 201306;
4. 农业农村部大洋渔业资源环境科学观测实验站 上海海洋大学 上海 201306;
5. 潍坊市畜牧业发展中心 山东潍坊 261041
2. Key Laboratory of Marine Ecological Monitoring and Restoration Technologies, MNRs, Shanghai 201306, China;
3. National Distant-water Fisheries Engineering Research Center, Shanghai Ocean University, Shanghai 201306, China;
4. Scientific Observing and Experimental Station of Oceanic Fishery Resources, Ministry of Agriculture and Rural Affairs, Shanghai 201306, China;
5. Weifang Animal Husbandry Development Center, Weifang 261041, China
鸢乌贼(Sthenoteuthis oualaniensis)属头足纲(Cephalopoda)、柔鱼科(Ommastrephidae)、鸢乌贼属(Sthenoteuthis), 为暖水性大洋性种类, 广泛分布于太平洋和印度洋的热带与亚热带海域(40°N~40°S) (叶守建等, 2014)。与其他柔鱼类相比, 鸢乌贼资源分布广泛, 但集中分布在印度洋北部和我国南海(Zuev et al, 1994; Ikeda et al, 1997), 中东太平洋赤道海域的鸢乌贼常作为兼捕对象(陆化杰等, 2014)。2005~2007年我国对印度洋北部鸢乌贼资源开展调查, 最高产量达5 000 t (Chen et al, 2008)。鸢乌贼生命周期短、生长快速(Estupiñán-Montaño et al, 2019), 繁殖能力强(Chen et al, 2007a; 陆化杰等, 2018), 是极具商业开发价值的经济头足类, 同时在分布海域生态系统中起着重要作用(Kojadinovic et al, 2007)。角质颚作为头足类摄食器官, 具有耐腐蚀、结构稳定、储存信息丰富等特点, 被广泛用于研究头足类渔业生态学及资源评估等(方舟等, 2014; 陈子越等, 2019a)。近年来相关学者利用角质颚微结构对柔鱼科其他种类的渔业生物学特性进行了研究(胡贯宇等, 2015; 李建华等, 2018; 林静远等, 2019; 方舟等, 2020; 陈子越等, 2021), 但是对年龄与生长的研究多选择利用耳石微结构的方法(陆化杰等, 2012a, 2020a; 王洪浩等, 2022), 利用角质颚微结构对不同气候年间西北印度洋鸢乌贼生长特性的研究尚未见报道, 鉴于此, 本文根据不同气候年间2019年(厄尔尼诺年份)和2020年(正常年份)相同月份(2~5月)中国灯光罩网渔船在西北印度洋采集的1 896尾鸢乌贼样本, 对不同气候年间鸢乌贼角质颚微结构及生长特性开展对比研究, 为后续利用角质颚研究印度洋鸢乌贼生活史及资源评估等提供科学依据。
1 材料与方法 1.1 样本来源样品采集时间为2019年、2020年2~5月, 采集海域为西北印度洋13°05′~17°45′N、59°06~63°50′E (图 1), 采集渔船为舟山宁泰远洋渔业公司所属的“欣海1223”号灯光罩网渔船, 渔船参数为: 船长51.2 m, 船宽8.3 m, 型深4.1 m, 总吨位557 t, 主机1 155 kW, 副机600 kW。由于采样点纬度、经度跨度均为5°以内, 且两年间大部分采样海域高度重合, 本研究中忽略采样点差异性对角质颚微结构和生长可能产生的影响。
 |
| 图 1 采样站点图 Fig. 1 Site of investigations and samples |
每天从每个站点渔获物中随机挑选鸢乌贼样本20~30尾, 共采集1 896尾。经过冷冻保存后运回实验室, 进行渔业生物学实验并提取完整角质颚, 利用角质颚微结构对其日龄进行鉴定(陈子越等, 2021), 共得到588个有效年龄数据(2019年290个, 2020年298个), 样本信息见表 1。
| 年份 | 性别 | 采样经度 | 采样纬度 | 样本数/尾 |
| 2019 | 雌性 | 62°41′E~63°38′E | 16°10′N~17°12′N | 573 |
| 雄性 | 62°56′E~63°35′E | 16°10′N~16°48′N | 393 | |
| 2020 | 雌性 | 59°38′E~63°32′E | 15°33′N~17°47′N | 520 |
| 雄性 | 63°36′E~63°42′E | 14°29′N~16°30′N | 410 |
研究表明, 鸢乌贼角质颚微结构的生长纹位于喙部矢状切面上(胡贯宇等, 2017; 陈子越等, 2021), 角质颚切片的制作包括4个步骤, 即切割、包埋、研磨和抛光, 使用切割工具从角质颚上颚喙尖端偏下部分沿头盖顶部中线方向切开直至后缘, 将喙端的半个矢状切面平整剪下, 放入长方形包埋槽中, 调整角度使矢状切面中线与包埋槽平行, 利用硬化剂和冷埋树脂调配一定比例的树脂溶液, 缓慢均匀导入包埋槽中, 用以固定包埋角质颚(陆化杰等, 2020a; 陈子越等, 2021)。静置硬化后, 在Stuers专业研磨机上依次使用目数为120、600、1200、2000、2500型号的水磨砂纸对其双面进行研磨, 在Olympus光学显微镜下观察到较为清晰的生长轮纹切面后, 再用氧化铝抛光粉溶液在水绒布上进行抛光, 直至观察到理想的日龄生长轮纹(陆化杰等, 2020a)。
1.3 轮纹计数将制备好的角质颚切片在连接charged coupled device (CCD)系统的Olympus光学显微镜100倍下观察拍照, 然后利用PhotoShop 24.0软件对同一样本角质颚切片的多张不同部位照片进行后期合成, 得到完整的角质颚微结构图片(陆化杰等, 2020a; 陈子越等, 2021)。每一个角质颚切片的日龄生长纹由3个实验员分别计数一次, 采用变异系数(coefficient of variation, Cv)检验计数的准确性, 计算公式如下:
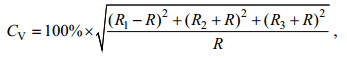 (1)
(1)式中, R为3人生长纹读数的均值, R1、R2、R3分别表示3人对同一切片样本的读数。
各自计数的日龄数与均值的差值低于5%即认为准确, 否则认定为无效, 取3次生长纹计数的均值为准确日龄(林静远等, 2019; 陈子越等, 2021)。
1.4 不同年间气候事件确定根据美国国家海洋和大气管理局定义, 海表温度距平指数连续3个月高于0.5 ℃为一次厄尔尼诺事件, 连续3个月低于0.5 ℃为一次拉尼娜事件(Chen et al, 2007b)。
2019年2~5月及之前海表温度距平指数均有连续3个月高于0.5 ℃, 认为发生厄尔尼诺事件; 2020年2~5月及之前均未有极端气候事件发生, 认为是正常月份(Chen et al, 2007b; 陆化杰等, 2018)。极端气候的出现, 可能会对鸢乌贼角质颚的生长生产影响(陆化杰等, 2018)。
1.5 数据处理(1) 利用典型相关分析法, 分析不同气候年间西北印度洋鸢乌贼角质颚各特征参数与日龄的相关性(李春喜等, 2008), 角质颚特征参数包括上头盖长(upper hood length, UHL)、上脊突长(upper crest length, UCL)、上侧壁长(upper lateral wall length, ULWL)、下脊突长(lower crest length, LCL)、下侧壁长(lower lateral wall length, LLWL)和下翼长(lower wing length, LWL) (何静茹等, 2020)。
(2) 利用协方差分析检验不同气候年间角质颚外部特征参数与日龄的生长关系是否存在显著性差异(陈子越等, 2019b; 何静茹等, 2020)。分别采用线性、指数、幂函数和对数函数方程, 在Excel 2019软件中对两年间鸢乌贼角质颚外部特征参数与日龄的生长关系进行拟合(陆化杰等, 2012b; 何静茹等, 2020)。应用赤池信息准则(Akaike’s information criterion, AIC)进行生长方程比较与选取, 取最小AIC值的模型为最适生长方程(陆化杰等, 2012b)。
对角质颚特征参数与日龄的关系进行线性回归分析(林静远等, 2019), 并对比不同气候年间角质颚生长差异性。
(4) 采用绝对生长率AGR (absolute growth rate, 单位: μm/d)和瞬时相对生长率IRGR (instantaneous relative growth rate, 单位: %/d)分析角质颚外形特征参数随日龄增长的生长变化情况(Chen et al, 2010; 陆化杰等, 2012b; 林静远等, 2019; 陈子越等, 2021), 公式分别为
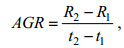 (2)
(2)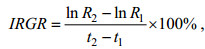 (3)
(3)式中, R2为t2日龄时角质颚特征参数长度(单位: μm); R1为t1日龄时角质颚特征参数长度(单位: μm); 本研究中日龄间隔为30 d, 即t2 – t2统一为30 d。
2 研究结果 2.1 角质颚微结构鸢乌贼角质颚上颚喙部截面RSS (rostrum sagittal section)微结构如图 2所示, 主要由头盖(位于背侧)和脊突(位于腹侧)两部分组成(图 2a), 角质颚上颚RSS微结构的日龄生长纹路明显, 以明、暗带交替的方式组成, 呈周期性生长(图 2b)。头盖和脊突两部分在内轴区域存在明显的分界线, 同时上下两部分的生长纹以“ < ”形相交于内轴, 这两部分生长纹宽度、数目及间隔均有不同, 头盖部的生长纹较为清晰, 喙端存在不同程度的磨损和缺刻, 喙顶端的生长纹间隔较窄, 同时同一条生长纹越靠近内轴宽度越宽(图 2c, 2d)。2019年(图 2e)明暗带均窄于2020年(图 2f), 每条生长纹间隔及宽度都较小, 色素沉积较2020年更浅, 轮纹暗带颜色更深, 喙端圆滑无缺刻且磨损较轻, 生长纹相对明显, 同时上颚RSS更易出现标记纹及一些纹路异常变化(图 2b)。
 |
| 图 2 鸢乌贼上角质颚微结构示意图 Fig. 2 The upper-beak microstructure of S. oualaniensis 注: a. 鸢乌贼上角质颚喙部; b. 头盖部生长纹及标记纹; c. 2019年鸢乌贼角质颚上颚喙部矢状切面微结构; d. 2020年鸢乌贼角质颚上颚喙部矢状切面微结构; e. 2019年明暗带及生长纹; f. 2020年明暗带及生长轮纹 |
协方差分析表明, 西北印度洋鸢乌贼日龄与UHL (F=8.479, P < 0.01)、UCL (F=32.251, P < 0.01)、ULWL (F=57.959, P < 0.01)、LCL (F=21.125, P < 0.01)、LLWL (F=38.009, P < 0.01)和LWL (F=14.131, P < 0.01) 6项特征参数均存在年间极显著差异。因此, 分年间研究角质颚外形特征参数与日龄的生长关系。典型相关分析表明, 两年间鸢乌贼日龄与角质颚各特征参数的相关性均达到了极显著水平(P < 0.01), 而且, 2019年日龄与角质颚各特征参数相关系数较2020年更高, 均达到0.85以上(表 2)。线性回归分析表明, 2020年角质颚特征参数与日龄拟合线性回归的a值比2019年的高, b值除LLWL外, 也均高于2019年, 详细结果见表 2。
| 特征参数 | 年份 | 回归分析 | 相关系数 | |||||
| 方差MS | F | a | b | R2 | P | |||
| UHL | 2019 | 2 364.461 | 527.226 | 0.112 | –7.228 | 0.763 | <0.01 | 0.873** |
| 2020 | 2 520.714 | 360.624 | 0.137 | 5.103 | 0.686 | <0.01 | 0.828** | |
| UCL | 2019 | 2 699.997 | 684.409 | 0.120 | –5.721 | 0.807 | <0.01 | 0.898** |
| 2020 | 4 082.048 | 388.361 | 0.174 | 4.151 | 0.702 | <0.01 | 0.838** | |
| ULWL | 2019 | 1 372.518 | 652.278 | 0.085 | –2.542 | 0.799 | <0.01 | 0.894* |
| 2020 | 2 886.398 | 360.304 | 0.147 | 2.584 | 0.686 | <0.01 | 0.828* | |
| LCL | 2019 | 723.865 | 536.709 | 0.062 | –3.558 | 0.766 | <0.01 | 0.875** |
| 2020 | 1 111.203 | 228.704 | 0.091 | –0.095 | 0.581 | <0.01 | 0.762** | |
| LLWL | 2019 | 1 772.104 | 461.283 | 0.097 | –5.703 | 0.738 | <0.01 | 0.859** |
| 2020 | 3 238.549 | 319.487 | 0.155 | –6.623 | 0.659 | <0.01 | 0.812** | |
| LWL | 2019 | 553.953 | 436.202 | 0.054 | –3.477 | 0.727 | <0.01 | 0.853** |
| 2020 | 681.571 | 353.180 | 0.071 | 1.364 | 0.682 | <0.01 | 0.826** | |
| 注: **表示极显著相关(P<0.01) | ||||||||
协方差分析表明, 2019年角质颚外形特征参数UHL (F=16.96, P < 0.01)、UCL (F=18.33, P < 0.01)、ULWL (F=13.19, P < 0.01)、LCL (F=26.73, P < 0.01)、LLWL (F=32.66, P < 0.01)和LWL (F=15.22, P < 0.01)与日龄的关系均存在性别间显著性差异, 因此分不同性别研究2019年样本角质颚外形特征参数与日龄的关系。
通过方程的拟合、优化和AIC的比较, 得到雌性个体所有外形特征参数与日龄的关系均最适用指数函数表示; 雄性个体除LCL与日龄的关系最适用指数函数表示外, 其余均最适用幂函数表示(图 3)。
 |
| 图 3 2019年鸢乌贼日龄与角质颚生长关系 Fig. 3 Relationship between age and beak of S. oualaniensis in 2019 |
协方差分析表明, 2020年角质颚外形特征参数UHL (F=61.34, P < 0.01)、UCL (F=84.88, P < 0.01)、ULWL (F=75.56, P < 0.01)、LCL (F=47.12, P < 0.01)、LLWL (F=74.27, P < 0.01)和LWL (F=57.62, P < 0.01)与日龄的关系均存在性别间显著性差异。因此, 分不同性别研究2020年样本角质颚外形特征参数与日龄的关系。
通过方程的拟合、优化和AIC的比较, 得到2020年雌性个体所有外形特征参数与日龄的关系均最适用对数函数表示, 雄性个体所有外形特征参数与日龄的关系均最适用线性函数表示(图 4)。
 |
| 图 4 2020年鸢乌贼日龄与角质颚生长关系 Fig. 4 Relationship between age and beak of S. oualaniensis in 2020 |
本研究表明, 不同年间, UHL、UCL、ULWL、LCL、LLWL和LWL均随日龄增加而增加, 其中除2020年雄性个体6项特征参数的AGR和IGRG随日龄增加呈现先下降后上升的“U”型趋势外, 其余特征参数的AGR和IGRG随日龄的增加都明显呈相反的倒“U”型趋势, 且生长率在不同年间的达到峰值时的日龄区间也不同(图 5, 图 6, 图 7, 图 8)。
 |
| 图 5 雌性鸢乌贼角质颚外部形态参数绝对生长率与日龄关系 Fig. 5 Relationship between AGR of morphological characteristic parameters and age for females beak |
 |
| 图 6 雌性鸢乌贼角质颚外部形态参数瞬时相对生长率与日龄关系 Fig. 6 Relationship between IRGR of morphological characteristic parameters and age for females beak |
 |
| 图 7 雄性鸢乌贼角质颚外部形态参数绝对生长率与日龄关系 Fig. 7 Relationship between AGR of morphological characteristic parameters and age for males' beak |
 |
| 图 8 雄性鸢乌贼角质颚外部形态参数瞬时相对生长率与日龄关系 Fig. 8 Relationship between IRGR of morphological characteristic parameters and age for males' beak |
对于雌性样本, 2019年样本日龄介于140~260 d之间, 各特征参数AGR呈明显平缓上升趋势, 231~260 d时各特征参数的AGR值均达到峰值, 之后逐渐下降, 其中ULWL的增长较为迅速(图 5)。2020年日龄介于140~170 d时, 各特征参数AGR呈明显上升趋势, 上升坡度较大; 171~200 d时, 各特征参数的AGR最大; 日龄大于200 d时, 各特征参数的AGR逐渐下降; 但在日龄 > 260 d时, UCL、ULWL、LCL的AGR又逐渐增大(图 5)。样本的IGRG在不同气候年间的变化趋势与AGR的变化保持基本一致, 其中, 2019年的UHL和LLWL在231~260 d时IGRG增长明显, 超过0.5%/d (图 6)。
对于雄性样本, 2019年角质颚各外形特征参数的生长率变化趋势不同, UHL、UCL和LWL日龄在140~170 d时AGR呈上升趋势, 171~200 d时AGR最大, 日龄大于200 d时AGR逐渐下降; ULWL、LLWL和LCL日龄在140~200 d时AGR呈上升趋势, 201~230 d区间内AGR最大, 日龄大于200 d时AGR逐渐下降(图 7)。2020年雄性样本日龄范围较小, 各个日龄分组间变化趋势较小, 呈先下降后上升的趋势, 在171~200 d时内AGR最小(图 7)。
总体而言, 雌性样本所有特征参数的AGR和UHL、LLWL的IGRG, 2020均大于2019年(图 5, 图 6); 雌性样本特征参数UCL、ULWL、LCL和LWL的IGRG, 以及雄性样本所有特征参数的AGR和IGRG, 2019均大于2020年(图 7, 图 8)。
3 讨论 3.1 角质颚微结构本研究表明, 与头足类耳石、内壳等硬组织一样, 西北印度洋鸢乌贼角质颚存在规律性明显的生长纹结构, 且头盖和脊突两部分纹路明显, 呈明、暗带交替的周期性生长, 这种生长纹的规律性可能是由于光-暗的周期性交替诱导产生(胡贯宇等, 2017)。头盖和脊突两部分生长纹以“ < ”形相交于内轴, 脊突部分生长纹排列紧密且多有重叠, 因此常以头盖部分排列有序的纹路计入生长纹。同时由于捕食过程中喙部尖端的磨损以及研磨过程中色素沉积的存在, 常选取喙部与尖端附近的背侧区域读取喙部生长纹, 这在柔鱼科其他头足类中也得到应用(陆化杰等, 2012b; 胡贯宇等, 2017; 陈子越等, 2021)。与柔鱼科其他头足类相似(Chen et al, 2010; Doubleday et al, 2011; 陈子越等, 2021), 西北印度洋鸢乌贼角质颚生长纹也保持“一日一纹”的生长规律。通过对比两年间鸢乌贼角质颚的生长纹发现, 2019年生长纹宽度及间隔都较2020年窄, 暗带颜色较深, 色素沉积也较浅, 这种现象可能是由于两年间样本的性腺成熟度不同导致, 陆化杰和陈炫妤分别对中国南海鸢乌贼和西北印度洋鸢乌贼角质颚色素沉积特性研究中也发现了类似情况(陈炫妤等, 2020; 陆化杰等, 2020b)。本研究通过对比两年样本组成发现, 2019年样本性腺发育主要处于未成熟阶段, 2020年样本则大部分已达到成熟阶段, 因此性腺成熟度可能是西北印度洋鸢乌贼角质颚生长纹颜色以及色素沉积差异的原因之一(陈炫妤等, 2020)。有研究表明, 鸢乌贼的摄食结构随个体的生长发生变化(Shchetinnikov, 1992; Kojadinovic et al, 2007; 陆化杰等, 2014), 个体越大, 角质颚撕碎食物的能力越强, 使用频率也越大, 因此, 个体较小的2019年样本, 食物需求量相对较小, 上颚RSS喙端圆滑少有缺刻, 磨损程度也较轻。此外, 2019年是厄尔尼诺年, 水文条件波动频繁, 表征生活史特殊事件(如产卵、洄游)或者外界环境变化(如水温波动、风暴或者捕食者的袭击)的标记纹也较多, 这与其他头足类的研究结果基本一致(Perales-Raya et al, 2010, 2014a, 2014b; 胡贯宇等, 2017)。
3.2 角质颚外部特征参数与日龄关系差异研究表明, 西北印度洋鸢乌贼角质颚特征参数与日龄的关系存在年间显著性差异, 且同一年份角质颚特征参数与日龄的关系均存在性别间显著性差异, 这与秘鲁外海茎柔鱼(Dosidicus gigas) (胡贯宇等, 2017)、西南大西洋阿根廷滑柔鱼(Illex argentinus) (陆化杰等, 2012b)的结果不完全相同, 可能是因为鸢乌贼不同阶段雌、雄个体摄食习性不同导致(何静茹等, 2020)。2019年雌性个体所有外形特征参数与日龄的关系均最适用指数函数表示, 而雄性个体的外部形态参数中除LCL最适用指数函数外, 其余均最适用幂函数表示; 2020年雌性个体所有外形特征参数与日龄的关系均最适用对数函数表示, 而雄性则均最适用线性函数表示。2020年雄性样本角质颚的生长特性与秘鲁外海茎柔鱼(胡贯宇等, 2017)、南海剑尖枪乌贼(Uroteuthis edulis) (林静远等, 2019)角质颚外形生长方程相同, 而与同属于柔鱼科的西南大西洋阿根廷滑柔鱼秋、冬季产卵群的角质颚生长方程不一致, 可能是由于样本属于不同的孵化群体造成的, 有研究表明不同的孵化群体角质颚外形特长特性不同(陆化杰等, 2012b)。研究表明, 2020年所有个体外形特征参数生长关系的生长指数(b值)均大于2019年, 说明2020年角质颚外形生长比2019年快。不同气候年间西北印度洋鸢乌贼生活史所经历的海洋环境不同, 食物种类丰度存在差异, 也会造成头足类角质颚的生长差异, 这与陈子越等(2021)和Keyl等(2011)的研究结果类似。
3.3 角质颚外部特征参数生长率研究表明, 不同气候年间西北印度洋鸢乌贼角质颚各特征参数的生长率存在差异, 2019年和2020年雌性样本绝对生长率AGR均呈现倒“U”型增长趋势, 其中2019年雌性在231~260 d时各特征参数的AGR值均达到峰值; 2019年雄性在171~200 d内AGR最大, ULWL、LLWL和LCL在201~230 d内AGR最大; 2020年雌性在171~200 d内, 各特征参数AGR最大; 2020年雄性变化趋势呈“U”型。鸢乌贼生长极易受到环境影响(何静茹等, 2020; 陈子越等, 2021), 由于2019年为厄尔尼诺年份, 水文条件波动频繁, 西北印度洋海域水温升高(陆化杰等, 2020c), 个体相对2020年小, 成活率以及摄食结构都会受到影响, 这与陆化杰等的研究结果一致(陆化杰等, 2021a)。本研究结果表明, 各特征参数随日龄的生长速率在不同气候年间存在差异, 可能是因为温度升高导致鸢乌贼个体减小, 雌雄间的个体差异变大, 因此角质颚各特征参数的生长与个体保持一致(何静茹等, 2021), 也存在差异。此外, 本研究观察到角质颚不同部位生长不同, 表征头盖部分和脊突部分的外形特征参数生长速率较表征侧壁部分和翼部的形态参数更快, 其中脊突部分生长速率最快, 翼部生长速率最慢, 而且上颚各外形特征参数生长速率大于下颚, 与其他学者关于北太平洋柔鱼(Ommastrephes bartramii) (方舟等, 2020)和鸢乌贼(何静茹等, 2020)的研究结果基本一致。角质颚外形生长存在差异的原因可以总结为4点: (1) 环境条件的突变: 角质颚生长过程中, 突变的水文条件使某一部分生长加速或变缓; (2) 肌肉转变: 有学者指出头足类在生长的过程中, 下颌肌肉会牵引角质颚发生一定程度的曲线旋转(Uyeno et al, 2005; 陆化杰等, 2014), 使得角质颚各个部位的生长速度存在不同; (3) 食性转变: 在鸢乌贼个体从幼稚鱼到成鱼的转变过程中, 捕食的种类也从浮游动物和甲壳类转变为以小型鱼类和头足类为主(Chen et al, 2007a; 陆化杰等, 2014), 还存在明显的自相残食现象, 个体撕碎食物的能力增强; (4) 角质颚各部分功能差异: 上颚是摄食功能的主体, 生长速度会大于下颚, 脊突部分和头盖部分主要起到支撑作用, 因此在个体发育的前期生长速率远远领先, 而在成体发育后, 喙部快速生长, 用以撕碎体型较大的食物(Jin et al, 2017)。同时最新研究表明, 头足类组织微塑料的沉积会影响到头中类的生长, 并最终影响到头足类角质颚的生长变化(陆化杰等, 2021b, 2022)。
4 结论不同气候年间西北印度洋鸢乌贼角质颚微结构存在差异性, 厄尔尼诺年份(2019年)明暗带均窄于正常年份(2020年), 生长纹间隔及宽度也相对较小, 色素沉积较浅, 轮纹暗带颜色更深, 生长纹相对明显, 同时上颚RSS更易出现标记纹及一些纹路异常变化。不同气候年间角质颚外部形态生长方程存在差异性, 厄尔尼诺年份(2019年)雌性个体所有外形特征参数与日龄的关系均最适用指数函数表示, 雄性个体除LCL与日龄的关系最适用指数函数表示外, 其余均最适用幂函数表示; 正常年份(2020年)雌性个体所有外形特征参数与日龄的关系均最适用对数函数表示, 雄性个体所有外形特征参数与日龄的关系均最适用线性函数表示。不同气候年间角质颚外形特殊参数生长率也存在差异性, 厄尔尼诺年份(2019年)雄性样本所有特征参数的绝对和相对生长率, 雌性样本的UCL、ULWL、LCL和LWL的相对生长率均大于正常年份(2020年), 但雌性样本的所有特征参数的绝对生长率和UHL、LLWL的相对生长率则呈现相反的态势。
作为短生命周期种类, 头足类的生长极易受到气候变化的影响。本研究选取厄尔尼诺年份和正常年份的西北印度洋鸢乌贼角质颚为研究对象, 通过研磨获取其日龄数据, 对比分析了不同气候年间的角质颚微结构、外形生长和生长率的差异性。然而, 气候变化的影响是多样、复杂的, 尤其西北印度洋还受到印度洋偶极子的影响, 多重因素的影响势必会导致西北印度洋的海温、盐度等海洋环境因子产生变化, 究竟哪些因子对西北印度洋鸢乌贼角质颚的生长变化起着主导作用, 后续还需要深入开展研究。同时由于采样难度所限(既要保证相同采样时间和海域, 又要符合不同气候年间要求), 本研究只采用了2019和2020年两年的数据, 后续团队将持续采样, 扩大时间和范围, 进一步完善相关研究。
王洪浩, 陆化杰, 何静茹, 等, 2022. 西北印度洋海域鸢乌贼耳石微结构及生长特性. 应用生态学报, 33(2): 3419-3426 |
方舟, 陈新军, 陆化杰, 等, 2014. 头足类角质颚研究进展Ⅰ——形态、结构与生长. 海洋渔业, 36(1): 78-89 |
方舟, 陈新军, 瞿俊跃, 等, 2020. 北太平洋柔鱼角质颚形态及生长年间差异. 上海海洋大学学报, 29(1): 109-120 |
叶守建, 周劲望, 杨铭霞, 等, 2014. 全球头足类资源开发现状分析及发展建议. 渔业信息与战略, 29(1): 11-17 |
李建华, 张鑫浩, 金岳, 等, 2018. 基于耳石和角质颚微结构的中国枪乌贼年龄与生长比较. 海洋渔业, 40(5): 513-521 DOI:10.3969/j.issn.1004-2490.2018.05.001 |
李春喜, 邵云, 姜丽娜, 2008. 生物统计学. 4版. 北京: 科学出版社, 143-160
|
何静茹, 陆化杰, 陈炫妤, 等, 2020. 冬春季西北印度洋鸢乌贼角质颚外部形态及生长特性. 应用生态学报, 31(8): 2775-2784 |
何静茹, 陆化杰, 陈炫妤, 等, 2021. 西北印度洋鸢乌贼角质颚外形变化的影响因素. 应用生态学报, 32(5): 1881-1889 |
陆化杰, 王从军, 陈新军, 2014. 4-6月东太平洋赤道公海鸢乌贼生物学特性初步研究. 上海海洋大学学报, 23(3): 441-447 |
陆化杰, 王洪浩, 刘凯, 等, 2020c. 厄尔尼诺发生期冬春季西北印度洋鸢乌贼耳石外形生长特性. 生态学杂志, 39(11): 3694-3703 |
陆化杰, 宁欣, 刘维, 等, 2021a. 不同气候条件下南海西沙海域鸢乌贼(Sthenoteuthis oualaniensis)渔业生物学比较研究. 海洋与湖沼, 52(4): 1029-1038 |
陆化杰, 刘凯, 陈子越, 等, 2022. 南海西沙群岛海域鸢乌贼Sthenoteuthis oualaniensis胃组织微塑料沉积特性研究. 海洋与湖沼, 53(1): 187-194 |
陆化杰, 刘凯, 欧玉哲, 等, 2021b. 微塑料染污及其对不同栖息地、不同食性海洋鱼类影响的研究进展. 水产学报, 45(12): 2099-2111 |
陆化杰, 张旭, 童玉和, 等, 2020a. 中国南海西沙群岛海域鸢乌贼耳石微结构及生长特性. 水产学报, 44(5): 767-776 |
陆化杰, 陈子越, 宁欣, 等, 2020b. 中国南海西沙群岛海域鸢乌贼角质颚色素沉积变化. 生态学杂志, 39(5): 1600-1608 |
陆化杰, 陈新军, 2012a. 利用耳石微结构研究西南大西洋阿根廷滑柔鱼的日龄、生长与种群结构. 水产学报, 36(7): 1049-1056 |
陆化杰, 陈新军, 方舟, 2012b. 西南大西洋阿根廷滑柔鱼2个不同产卵群间角质颚外形生长特性比较. 中国海洋大学学报, 42(10): 33-40 |
陆化杰, 童玉和, 刘维, 等, 2018. 厄尔尼诺年春季中国南海中沙群岛海域鸢乌贼的渔业生物学特性. 水产学报, 42(6): 912-921 |
陈子越, 陆化杰, 童玉和, 等, 2019a. 个体差异对西沙群岛海域鸢乌贼角质颚外部形态变化的影响. 水产学报, 43(12): 2501-2510 |
陈子越, 陆化杰, 童玉和, 等, 2019b. 中国南海西沙群岛海域鸢乌贼角质颚生长特性. 上海海洋大学学报, 28(3): 373-383 |
陈子越, 武孝慈, 陆化杰, 等, 2021. 南海西沙群岛海域鸢乌贼(Sthenoteuthis oualaniensis)角质颚微结构及生长特性研究. 海洋与湖沼, 52(5): 1293-1302 |
陈炫妤, 陆化杰, 王洪浩, 等, 2020. 西北印度洋鸢乌贼角质颚色素沉积特性分析. 动物学杂志, 55(4): 468-476 |
林静远, 刘必林, 王立权, 2019. 基于角质颚微结构的剑尖枪乌贼的日龄与生长. 华东理工大学学报(自然科学版), 45(5): 775-782 |
胡贯宇, 陈新军, 刘必林, 等, 2015. 茎柔鱼耳石和角质颚微结构及轮纹判读. 水产学报, 39(3): 361-370 |
胡贯宇, 金岳, 陈新军, 2017. 秘鲁外海茎柔鱼角质颚的形态特征及其与个体大小和日龄的关系. 海洋渔业, 39(4): 361-371 |
CHEN X J, LIU B L, CHEN Y, 2008. A review of the development of Chinese distant-water squid jigging fisheries. Fisheries Research, 89(3): 211-221 |
CHEN X J, LIU B L, TIAN S Q, et al, 2007a. Fishery biology of purpleback squid, Sthenoteuthis oualaniensis, in the northwest Indian Ocean. Fisheries Research, 83(1): 98-104 |
CHEN X J, LU H J, LIU B L, et al, 2010. Age, growth and population structure of jumbo flying squid, Dosidicus gigas, based on statolith microstructure off the Exclusive Economic Zone of Chilean waters. Journal of the Marine Biological Association of the United Kingdom, 91(1): 229-235 |
CHEN X J, ZHAO X H, CHEN Y, 2007b. Influence of El Niño/La Niña on the western winter–spring cohort of neon flying squid (Ommastrephes bartramii) in the northwestern Pacific Ocean. ICES Journal of Marine Science, 64(6): 1152-1160 |
DOUBLEDAY Z A, WHITE J, PECL G T, et al, 2011. Age determination in merobenthic octopuses using stylet increment analysis: assessing future challenges using Macroctopus maorum as a model. ICES Journal of Marine Science, 68(10): 2059-2063 |
ESTUPIÑÁN-MONTAÑO C, CEDEÑO-FIGUEROA L, ESTUPIÑÁN-ORTIZ J F, et al, 2019. Feeding habits and trophic level of the smooth hammerhead shark, Sphyrna zygaena (Carcharhiniformes: Sphyrnidae), off Ecuador. Journal of the Marine Biological Association of the United Kingdom, 99(3): 673-680 |
IKEDA Y, ARAI N, SAKAMOTO W, et al, 1997. Comparison on trace elements in squid statoliths of different species' origin: as available key for taxonomic and phylogenetic study. International Journal of PIXE, 7(3/4): 141-146 |
JIN Y, LIU B L, LI J H, et al, 2017. Identification of three common Loliginidae squid species in the South China Sea by analyzing hard tissues with geometric outline method. Journal of Ocean University of China, 16(5): 840-846 |
KEYL F, ARGÜELLES J, TAFUR R, 2011. Interannual variability in size structure, age, and growth of jumbo squid (Dosidicus gigas) assessed by modal progression analysis. ICES Journal of Marine Science, 68(3): 507-518 |
KOJADINOVIC J, CORRE M L, COSSON R P, et al, 2007. Trace elements in three marine birds breeding on reunion island (western Indian Ocean): Part 1—Factors influencing their bioaccumulation. Archives of Environmental Contamination and Toxicology, 52(3): 418-430 |
PERALES-RAYA C, ALMANSA E, BARTOLOMÉ A, et al, 2014a. Age validation in Octopus vulgaris beaks across the full ontogenetic range: beaks as recorders of life events in octopuses. Journal of Shellfish Research, 33(2): 481-493 |
PERALES-RAYA C, BARTOLOMÉ A, GARCÍA-SANTAMARÍA M T, et al, 2010. Age estimation obtained from analysis of octopus (Octopus vulgaris Cuvier. 1797) beaks: Improvements and comparisons. Fisheries Research, 106(2): 171-176 |
PERALES-RAYA C, JURADO-RUZAFA A, BARTOLOMÉ A, et al, 2014b. Age of spent Octopus vulgaris and stress mark analysis using beaks of wild individuals. Hydrobiologia, 725(1): 105-114 |
SHCHETINNIKOV A S, 1992. Feeding spectrum of squid Sthenoteuthis oualaniensis (OEGOPSIDA) in the eastern Pacific. Journal of the Marine Biological Association of the United Kingdom, 72(4): 849-860 |
UYENO T A, KIER W M, 2005. Functional morphology of the cephalopod buccal mass: a novel joint type. Journal of Morphology, 264(2): 211-222 |
ZUEV G V, NIKOLSKY V N, SHESALIN M V, 1994. The biology and resources of the purpleblack flying squid (Sthenoteuthis oualaniensis) in the Arabian Sea. Biological Processes in the Arabian Sea: 167-172 |