 2023, Vol. 54
2023, Vol. 54

海州湾中国毛虾(Acetes chinensis)种群生物学特征和资源开发状态研究

http://dx.doi.org/10.11693/hyhz20220600160
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 吴晓睿, 宋大德, 熊瑛, 仲霞铭, 李纲, 杨帆, 康中杰, 李国东, 李冬佳, 施金金, 闫欣. 2023.
- WU Xiao-Rui, SONG Da-De, XIONG Ying, ZHONG Xia-Ming, LI Gang, YANG Fan, KANG Zhong-Jie, LI Guo-Dong, LI Dong-Jia, SHI Jin-Jin, YAN Xin. 2023.
- 海州湾中国毛虾(Acetes chinensis)种群生物学特征和资源开发状态研究
- POPULATION BIOLOGICAL CHARACTERISTICS AND EXPLOITATION STATUS OF ACETES CHINENSIS IN HAIZHOU BAY
- 海洋与湖沼, 54(2): 573-582
- Oceanologia et Limnologia Sinica, 54(2): 573-582.
- http://dx.doi.org/10.11693/hyhz20220600160
文章历史
-
收稿日期:2022-06-19
收修改稿日期:2022-08-07
引用本文
吴晓睿, 宋大德, 熊瑛, 仲霞铭, 李纲, 杨帆, 康中杰, 李国东, 李冬佳, 施金金, 闫欣. 2023. 海州湾中国毛虾(Acetes chinensis)种群生物学特征和资源开发状态研究[J]. 海洋与湖沼, 54(2): 573-582

WU Xiao-Rui, SONG Da-De, XIONG Ying, ZHONG Xia-Ming, LI Gang, YANG Fan, KANG Zhong-Jie, LI Guo-Dong, LI Dong-Jia, SHI Jin-Jin, YAN Xin. 2023. POPULATION BIOLOGICAL CHARACTERISTICS AND EXPLOITATION STATUS OF ACETES CHINENSIS IN HAIZHOU BAY[J]. Oceanologia et Limnologia Sinica, 54(2): 573-582.
海州湾中国毛虾(Acetes chinensis)种群生物学特征和资源开发状态研究
1. 上海海洋大学海洋科学学院 上海 201306;
2. 江苏省海洋水产研究所 江苏南通 226007;
3. 上海海洋大学水产与生命学院 上海 201306
2. 江苏省海洋水产研究所 江苏南通 226007;
3. 上海海洋大学水产与生命学院 上海 201306
摘要:为掌握我国限额捕捞物种海州湾中国毛虾(Acetes chinensis)的资源动态, 于2020、2021年6~7月分别采集了1 094、1 700尾中国毛虾样本, 测定其体长、体质量和性别等特征, 并运用FiSAT Ⅱ软件对生长、死亡参数及资源利用水平等进行评估。结果显示: 2020、2021年限额捕捞期间其平均体长分别为32.30、35.73 mm, 雌雄性比分别为1.52:1、1.62:1, 体长大于35 mm的毛虾中雌性比例分别为98.5%、90.0%, 并得出了体质量-体长关系式; 2020、2021年自然死亡系数(M)分别为0.55、0.61 month–1, 捕捞死亡系数(F)分别为0.62、0.27 month–1; 2020、2021年开捕体长(Lc)分别为26.28、29.73 mm, 均小于最适开捕体长(Lopt) 33 mm; 2020、2021年资源开发率(E)分别为0.53、0.31, 均小于当年E0.1 (0.60、0.77), 表明海州湾中国毛虾资源尚未过度捕捞。鉴于我国实施伏季休渔期毛虾限额捕捞的规模不断扩大, 应谨慎增加捕捞努力量, 将开发率E有效控制在E0.1以内。
关键词:中国毛虾 海州湾 体长股分析法 群体生物学特征 开发率
POPULATION BIOLOGICAL CHARACTERISTICS AND EXPLOITATION STATUS OF ACETES CHINENSIS IN HAIZHOU BAY
1. College of Marine Sciences of Shanghai Ocean University, Shanghai 201306, China;
2. Jiangsu Marine Fisheries Research Institute, Nantong 226007, China;
3. College of Fisheries and Life Science of Shanghai Ocean University, Shanghai 201306, China
2. Jiangsu Marine Fisheries Research Institute, Nantong 226007, China;
3. College of Fisheries and Life Science of Shanghai Ocean University, Shanghai 201306, China
Abstract: Acetes chinensis is a fishery resource of great economic importance in China, Republic of Korea, and Japan. However, little is known about its resource assessment and population biological characteristics especially in China waters from the early 2000s to present. In 2020 and 2021, A. chinensis boost the local fishery economy in Haizhou Bay in the short run. Therefore, taking the opportunity of A. chinensis in Haizhou Bay under the pilot of marine fisheries TAC (Total Allowable Catch), 1 094 and 1 700 fish specimens were sampled in 2020 and 2021, respectively, from the special fishing planning area of A. chinensis in the Haizhou Bay. The population parameters of A. chinensis were estimated by using the Length based Cohort Analysis (LCA) in FiSAT II software. Results show that the average body length of A. chinensis in 2020 and 2021 were 32.30 and 35.73 mm, respectively; sex ratio of A. chinensis in 2020 and 2021 were 1.52:1 and 1.62:1, respectively; Females of the A. chinensis with body length over 35 mm in these two years were 98.5% and 90.0%, respectively. And the relational equation between body length and body weight has been solved. Nature mortality coefficient (M) and fishing mortality coefficient (F) were 0.55 and 0.62 month–1 in 2020, respectively, while they were 0.61 and 0.27 month–1 in 2021, respectively. The catchable length of A. chinensis in 2020 and 2021 were 26.28 and 29.73 mm, respectively, which are both smaller than the optimum ca tchable length (33 mm). In 2020 and 2021, exploitation rate (E) of A. chinensis resources in Haizhou Bay were 0.53 and 0.31, respectively, which were lower than E0.1 in corresponding year, indicating minimal fishing effects. In view of the expansion of the scale of marine fisheries TACs in China, fishing effort should be increased carefully, and the exploitation rate shall be effectively controlled by the E0.1 to achieve the optimum state of conservation and utilization.
Key words:
Acetes chinensis Haizhou Bay Length based Cohort Analysis (LCA) population biological characteristics exploitation rate
中国毛虾(Acetes chinensis) (以下简称“毛虾”)属软甲纲(Malacostraca)、十足目(Decapoda)、樱虾科(Sergestidae)、毛虾属(Acetes), 为小型、透明、浮游型虾类, 广泛分布于西北太平洋沿岸近海水域(宋海棠等, 2012)。毛虾在我国近海具有较高产量, 其中在渤海湾、莱州湾毛虾年产量一般为4~5万t, 最高可达8万t以上, 约占整个渤海区毛虾总产量的2/3 (张孟海, 1992)。毛虾不仅在我国近海产量较高, 1995~2000年韩国年均捕捞中国毛虾和日本毛虾(Acetes japonicus)约1.66万t, 占其虾类产量40%以上(Oh et al, 2003)。我国江苏海域海州湾百年来盛产高品质毛虾, 作为高价值海产品常出口韩国、日本和美国等国家; 2020年, 我国首次在江苏省连云港实施了毛虾限额捕捞试点, 6月15日~7月15日一个月实现相关产业联动效益约3亿元(江苏省人民政府, 2020), 近两年海州湾中国毛虾因其短期内高额经济效益而受到高度重视。此外, 毛虾作为关键初级消费者和饵料生物(郭斌等, 2010; 贺舟挺等, 2012; 王毅波等, 2019), 是食物网物质循环和能量流动的关键调节者, 其数量波动会对其他物种甚至整个生态系统产生深远影响(徐从军等, 2019, 2021)。基于毛虾在渔业经济和生态系统的重要性(陈立婧等, 2022), 亟需清晰完整地认知毛虾资源状态以期科学合理地利用该资源。
我国有关毛虾的研究始于20世纪50年代, 主要针对渤海海域开展了调查研究(刘瑞玉, 1956; 张孟海, 1992; 陈立婧等, 2022), 迄今为止, 海州湾毛虾的生物学研究尚未见报道。为此, 本文以2020年和2021年海州湾毛虾为研究对象, 通过分析其体长分布组成、体质量-体长关系来评估种群参数, 以期掌握目前海州湾毛虾生长特性及其资源利用现状, 旨在为其资源合理利用和保护提供数据支撑, 从而可持续地推进海州湾毛虾限捕捕捞试点工作。
1 材料与方法 1.1 样品来源于2020、2021年农业农村部规定的毛虾限额捕捞定点上岸处——江苏海州湾海域燕尾港码头, 采样区域见图 1。每天采样2~3次, 每次随机采集湿毛虾5 kg。
 |
| 图 1 2020年和2021年海州湾毛虾采样区域图 Fig. 1 The sampling regions of A. chinensis in the Haizhou Bay in 2020 and 2021 |
文中用于估算其自然死亡系数时所需的栖息地水域海洋表面温度来自于卫星遥感SST (sea surface temperature)数据, 其时间范围为2020年1月~2021年12月, 空间分辨率为1°, 时间分辨率为1月, 具体下载地址为: http://www.ocean.iap.ac.cn/pages/dataService/dataService.html?navAnchor=dataService。
1.2 研究方法 1.2.1 虾体测量从每次采集的样品中随机取50尾毛虾个体(采样信息详见表 1), 测量其体长(body length, 单位: mm)、体质量(body weight, 单位: g)并鉴别性别, 体长和体质量分别精确至0.1 mm和0.1 g。体长测量依据Mauchline(1980)的标准, 即由额角最前端至尾节末端之间的长度。
表 1 2020年和2021年海州湾毛虾采样信息及捕捞状况 Tab. 1 Sampling and fishing information of A. chinensis from the Haizhou Bay in 2020 and 2021
| 年份 | 月份 | 样本量/尾 | 捕捞渔船数/艘 |
| 2020 | 6月、7月 | 1 094 | 86 |
| 2021 | 6月、7月 | 1 700 | 33 |
2020、2021年毛虾捕捞期间渔获量估算参照李国东等(2021)方法, 该方法借助船位监控系统(vessel monitoring system, VMS)渔业大数据解译, 在一定程度上有效解决了我国近海渔捞日志空缺及真实性不足的问题(张胜茂等, 2014)。
1.2.3 生长方程及性比海州湾毛虾体质量与体长回归方程W = aLb由幂函数拟合, 式中W、L、a和b分别为体质量(单位: g)、体长(单位: mm)、条件因子和幂指数系数。
本研究基于海州湾毛虾体长分布范围特点, 以5 mm为间隔, 划分为20~25、26~30、……、51~55 mm的7个体长组, 分别探究其性比。
1.2.4 生长参数鉴于毛虾属短生命周期型虾类, 因此其寿命单位以月龄(Monthly age)表示(Rahman et al, 2018; 陈立婧等, 2022)。本研究中的毛虾体长以2 mm为组距, 2020、2021年毛虾体长组分别划分为14、16组, 在FiSAT Ⅱ (FAO ICLARM Stock Assessment Tools Ⅱ)软件中按照要求创建体长频率分布表, 利用FiSAT Ⅱ软件Assess模块的ELEFAN Ⅰ程序估算毛虾的生长系数(K)和渐近体长(L∞)。参数L∞和K值是依据拟合优度Rn估算的, Rn值用来确定“最佳”拟合体长频数数据的分布曲线, 具有最高Rn值的L∞和K值被认为是“最佳”估计。然后利用Pauly(1983)经验公式推算理论生长初始月龄(t0):
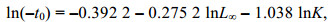 (1)
(1)式中, t0表示理论生长初始月龄, K表示生长系数, L∞表示渐近体长。
利用FiSAT Ⅱ软件中的Assess模块的体长变换渔获物曲线(length-converted catch curve)程序估算总死亡系数(Z), 公式:
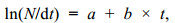 (2)
(2)式中, N为各体长组的尾数占总渔获尾数的百分比, dt为相应体长组的下限生长到上限所需的时间, a和b为常数, t为各体长组中值所对应的年龄, 总死亡系数即为下降部分的点做回归所得斜率的负值, 即Z = –b。
采用Pauly经验公式估算自然死亡系数(M) (Pauly, 1990), 公式:
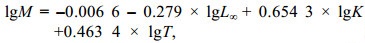 (3)
(3)式中, T为物种栖息水域的年平均表层水温, 2020年和2021年海州湾海域SST分别为17、17.2 ℃。
捕捞死亡系数F计算式为
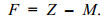 (4)
(4)渔业开发率E计算式为
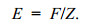 (5e)
(5e)最适开捕体长Lopt计算公式为
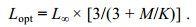 (6)
(6)通过线性长度转换渔获量曲线的左推法来确定捕获概率, 并利用Logit函数从渔获量曲线分析中得到捕获概率为25%、50%和75%时的选择体长L25、L50和L75, 并以L50为开捕体长(Lc)。
1.2.5 单位补充量模型分析Beverton等(1957)提出动态综合模型(dynamic pooled model), 也称Beverton- Holt (B-H)稳定状态模型, 用以解释鱼类种群参数和渔获量的关系, 渔获量(Y)以及单位补充量渔获量(Y′/R)的变化情况随着模型中的参数变化而变化。Pauly(1983)利用动态综合模性来预测Y′/R和B′/R (相对单位补充量生物量), 并验证了模型的可行性。Y′/R和B′/R曲线基于刀刃式选择假设模型(knife-edge model)建立, 根据计算的M/K和Lc/L∞值导出生物学参考点E。FiSAT Ⅱ软件中开发率(E)一般有下述几种水平: Emax、E0.1和E0.5, 分别表示为获得最大渔获量时的开发率、Y′/R边际增长减少10.0%时的开发率和资源量下降到原始水平50.0%时的开发率。
2 结果 2.1 体长分布2020、2021年分别于海州湾毛虾限额捕捞区域采集了毛虾1 094、1 700尾, 其体长范围分别为21~51、23~48 mm, 其中2020年和2021年雌性毛虾优势体长组范围分别为25~45、25~45 mm, 雄性毛虾优势体长组范围分别为21~35、25~40 mm。雄性在较小的体型等级中数量众多, 而雌性在较大的体型等级中占主要部分(图 2), t-检验结果均表明, 雌性和雄性之间的大小频数分布差异显著(P < 0.05)。
 |
| 图 2 2020年和2021年海州湾雌、雄毛虾体长频数分布及其体长正态分布函数曲线图 Fig. 2 Length-frequency distribution of males and females and the normal distribution curve of A. chinensis in the Haizhou Bay in 2020 and 2021 注: 实线曲线表示体长数据的正态分布曲线 |
2020年和2021年毛虾雌雄性比分别为1.52:1、1.62:1, 雌性比例均呈现随着体长的增大而增加的变化趋势。2020年海州湾毛虾雌性比例以体长30 mm为分界点显著增加(P < 0.05), 2021年雌性比例则以体长35 mm为分界点显著增加(P < 0.05) (图 3), 且2020和2021年体长超过35 mm的毛虾个体中雌性比例分别为98.5%、90.0%。
 |
| 图 3 2020年和2021年海州湾不同体长区间的毛虾性比图(雌性/总体) Fig. 3 Sex ratio (female no./total no.) of A. chinensis in the Haizhou Bay plotted at different intervals of body length in 2020 and 2021 注: 图中纵坐标值越大表示, 雌性毛虾占比越高; 纵坐标值越小, 雄性毛虾占比越高 |
渔获量估算参照李国东等(2021)方法, 2020、2021年毛虾捕捞期间毛虾渔获量分别为0.32、0.12万t。
2.3 体质量和体长关系2020年海州湾毛虾平均体长和平均体质量分别为(32.30±5.33) mm、(0.29±0.15) g, 其雌、雄性个体平均体长分别为(35.37±4.61) mm、(27.76±2.23) mm; 2021年海州湾毛虾平均体长和平均体质量分别为(35.73±5.40) mm、(0.30±0.14) g, 其雌、雄性个体平均体长分别为(38.50±4.83) mm、(31.15±2.41) mm。海州湾毛虾体质量和体长的关系呈幂函数相关(图 4), 2020年和2021年的体质量-体长关系式如下:
 |
| 图 4 2020年和2021年海州湾毛虾体质量与体长关系 Fig. 4 The length-weight relationships of A. chinensis in the Haizhou Bay in 2020 and 2021 |
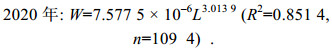 (6)
(6)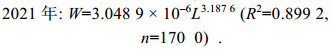 (7)
(7)t-检验结果表明, 2020年和2021年间的体质量-体长关系方程中的b值存在显著差异(P < 0.01)。
2.4 生长参数通过FiSAT Ⅱ软件中ELEEAN Ⅰ程序中的K扫描模块计算2020、2021年海州湾毛虾L∞分别为53.55和51.45 mm、K值分别为0.3和0.34, 根据Pauly提出的理论生长起点月龄的经验公式计算出计算2020、2021年t0分别为–0.79、–0.70 month。因此2020、2021年其Von Bertalanffy体长生长方程为
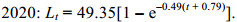 (8)
(8)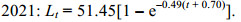 (9)
(9)利用体长变换渔获物曲线法对海州湾毛虾总死亡系数(Z)进行估算(图 5), 回归曲线斜率的绝对值即为Z, 2020、2021年估算得到的Z值分别为1.17、0.88 month–1, 因此F分别为0.62、0.27 month–1, E分别为0.53、0.31 (表 2)。
 |
| 图 5 海州湾毛虾体长变换渔获物曲线 Fig. 5 The curve of length-converted catch of A. chinensis in the Haizhou Bay in 2020 and 2021 注: 拟合过程中舍去了未达到补充年龄的黄点, 仅黑点被考虑用于计算总死亡系数 |
表 2 2020年和2021年海州湾毛虾种群生物学参数 Tab. 2 Parameters of the population biology of A. chinensis in the Haizhou Bay in 2020 and 2021
| 参数 | 估计值 | 参数 | 估计值 | |||
| 2020 | 2021 | 2020 | 2021 | |||
| L∞ | 53.55 mm | 51.45 mm | E0.1 | 0.60 | 0.77 | |
| K | 0.3 month–1 | 0.34 month–1 | E0.5 | 0.36 | 0.39 | |
| t0 | –0.79 month | –0.70 month | Emax | 0.75 | 0.90 | |
| Z | 1.17 month–1 | 0.88 month–1 | Lc(L50) | 26.28 mm | 29.73 mm | |
| M | 0.55 month–1 | 0.61 month–1 | L25 | 25.28 mm | 28.43 mm | |
| F | 0.62 month–1 | 0.27 month–1 | L75 | 27.27 mm | 31.03 mm | |
| E | 0.53 | 0.31 | Lopt | 33.26 mm | 32.22 mm | |
| M/K | 1.83 | 1.79 | ||||
海州湾毛虾的捕捞概率如图 6所示, 累计率至50%时, 2020年和2021年对应的Lc分别为26.28和29.73 mm (表 2)。
 |
| 图 6 2020年和2021年海州湾毛虾捕获概率的Logistic选择曲线 Fig. 6 Curve of logistic selection for probability of capture of A. chinensis in the Haizhou Bay |
采用FiSAT Ⅱ软件分析海州湾2020、2021年毛虾的开发率(E)与相对单位补充量渔获量(Y′/R)的关系(图 7)。其中2020年, M/K=1.83, Lc/L∞=0.49, 估算出Emax、E0.5、E0.1分别为0.75、0.36、0.60; 2021年, M/K=1.79, Lc/L∞=0.58, 估算出Emax、E0.5、E0.1分别为0.90、0.39、0.77。2020、2021年开发率水平分别为0.53、0.31, 均低于相应年份的最适开发率(Emax)。2020、2021年最适开捕体长Lopt分别为33.26、32.22 mm, 均大于相应年份的平均选择体长Lc。基于相对单位补充量等值曲线, 讨论E与Lc和Y’/R之间的动态变化趋势(图 8)。图 8深蓝区域至红色区域的渐进变化表示相对单位补充量的数值由低到高逐渐变大, 表明2020、2021年开发率A点均未达到红色区域内B点所指的最佳开发率, 意味着2020年和2021年海州湾毛虾资源尚未过度开发。
 |
| 图 7 2020年和2021年海州湾毛虾种群相对补充量渔获量、相对补充量生物量曲线 Fig. 7 Curves of relative yield-per-recruit (Y′/R) and relative biomass-per-recruit (B′/R) of A. chinensis in the Haizhou Bay in 2020 and 2021 注: 黄线、绿色和红色虚线分别表示Emax、E0.1和E0.5时的相对渔获量 |
 |
| 图 8 海州湾毛虾开发率和开捕体长与相对单位补充量的关系 Fig. 8 Relationships of relative yield-per-recruit (Y′/R) to exploitation ratio (E), and to the standard length at first catch (Lc) of A. chinensis in the Haizhou Bay in 2020 and 2021 |
近两年, 海州湾毛虾因其相当高的经济价值而备受关注。本文梳理了从北至南不同海域的毛虾个体体长规律以分析海州湾毛虾的关键特征。选取各海域繁殖期毛虾个体且体长最大的月份的数据进行论证: 1983~1985年历年2~11月份渤海湾和莱州湾, 毛虾6月中下旬体长平均值最大, 为25.74 mm, 优势体长组范围20~30 mm (张孟海, 1986); 2004~2006年渤海湾和莱州湾海域, 5月毛虾体长平均值最大, 为24.56 mm (曾现英等, 2016); 1982~1985年历年浙江北部沿岸海域, 5~7月毛虾个体最大, 优势体长组为27~35 mm (李星颉等, 1986); 2020、2021年6~7月海州湾毛虾体长平均值分别32.30、35.73 mm, 优势体长组分别为27~37和29~43 mm (图 2)。通过对比渤海湾、莱州湾、浙江北部近海的平均体长和优势体长组, 表明海州湾毛虾繁殖个体最大。尽管海州湾毛虾与上述毛虾来自不同年代、不同海域, 但从辽东湾1956~1967年3月的毛虾平均体长呈现下降趋势(陈俅, 1981)和渤海湾、莱州湾海域2004~2006年个体要小于1983~1985年的结果, 说明同一海域毛虾个体呈现出小型化趋势, 而2020~2021年海州湾毛虾个体仍大于20世纪80年代其他海域的个体, 因此, 可以得出“我国海域毛虾6~7月繁殖期, 海州湾毛虾繁殖个体最大”的结论。此外, 体质量-体长关系式中幂指数b值也侧面支持这一结论。2020、2021年海州湾毛虾体质量与体长关系式中的幂指数b值分别为3.01、3.19, 均在正常取值范围2.5~3.5之间(韩耀全等, 2017)。Oh等(2002)查明2000年5月~2001年10月间韩国西南部海域毛虾雌性和雄性个体生长方程b值分别为2.90、3.01。1998年8、10月和1999年5月黄河口水域和莱州湾毛虾的生物学特征数据显示, 所调查月份的雌性和雄性b值均小于2.5 (仲崇峻等, 2001), 远低于3。
究其原因, 生物内在因子(如基因)和外在因子(如温度、盐度和食物等)分别作用于鱼、虾类生长发育的代谢类型和代谢强度, 进而影响其生长进程(殷名称, 1995)。幂指数b值的差异也被认为与种群、水域、性别和年份等因素有关(詹秉义, 1995; 韩耀全等, 2017)。就海州湾毛虾而言, 生物内在因子(如基因)或不同世代对其个体大小的影响有待进一步验证。Wootton(1973)指出, 即使同一物种, 栖息生境的差异也可能导致鱼、虾类生长特征的不同。毛虾繁衍生长所需的主要饵料是浮游动植物, 而海水中的营养盐和叶绿素a是表征浮游动植物丰度的重要指标(王毅波等, 2019)。海州湾海岸线曲折呈凹形, 水深不超过30 m (Liu et al, 2013), 相较于其他海域营养盐和叶绿素a含量较高(Song et al, 2021)。Song等(2021)在分析山东半岛近海叶绿素a浓度时空变化发现, 海州湾夏季大面积地区的叶绿素浓度有明显增高的趋势, 而其他海域则相反。夏季, 黄海暖流和黄海沿岸流在海州湾海域交汇, 形成明显的上升流(Wei et al, 2020), 并且随着近年海州湾大量人工鱼礁区的建成, 鱼礁群周围也会产生明显的上升流和背涡流(罗文强等, 2021), 能从海底携带大量营养盐到表层, 一定程度上增加了营养盐的含量。同时, 夏季是丰水期, 大量的陆地径流向海州湾源源不断地输送有机物和营养盐(杨晓改, 2015; Song et al, 2021), 海州湾水域毛虾个体大可能与该海域丰富的营养盐和叶绿素a密切相关。
2020、2021年海州湾毛虾样本中雌雄性比均高于1.5且雌虾个体明显大于雄虾, 这与仲崇峻等(2001)探究莱州湾和黄河口水域繁殖期的毛虾雌雄性比和个体结论一致, 这可能与雌性毛虾季节性产卵迁移的因素有关(施仁德, 1986)。繁殖季节雌雄性比高于非繁殖季节的状况也与雌、雄虾的性成熟情况和生长速度有关(徐君义, 1986), 合适的性别比例是有助于雌雄交配和性腺发育(Claudia et al, 2004)。毛虾特许捕捞期间正处于毛虾产卵盛期, 其繁殖季节雌多雄少现象与葛氏长臂虾(Palaemon gravieri) (汤晓建等, 2019)和脊尾白虾(Exopalaemon carinicauda) (徐君义, 1986)生殖期间雌雄比特征一致。此外, 捕捞网具的选择性使得所使用的网具对大个体的雌性毛虾选择性要高, 因此也可能造成毛虾渔获中雌性个体数量高于雄性(Oh et al, 2003)。
3.2 毛虾资源开发状态2020、2021年海州湾毛虾生长系数K分别为0.3、0.34 month–1, 韩国西部近岸海域的毛虾K值为0.64 month–1 (Oh et al, 2003), 两个海域生长系数差异较大的原因除了与海域、年份相关外, 还有可能是Oh等(2003)数据来源为全年12个月的毛虾数据, 而本文采用的是海州湾毛虾繁殖期数据。毛虾繁殖期低K值主要反映了繁殖活动期间体内能量的消耗分解。2020、2021年海州湾毛虾自然死亡系数M分别为0.55、0.61 month–1, 捕捞死亡系数F分别0.62、0.27 month–1, 相较于M值, 2020年和2021年F值差异较大, 这与2020、2021年实际投入毛虾生产的捕捞努力量的差异相吻合(表 1)。2020、2021年海州湾毛虾开发率E分别为0.53、0.31, 均小于相应年份的E0.1、Emax值(表 2), 表明2020、2021年海州湾毛虾资源均未处于过度开发状态。就开发率而言, Emax被视为短生命种类的“目标”和“极限”参考点, Hoggarth等(2006)建议, 由于E0.1比Emax更谨慎、更适用, 应以其作为目标参考点。伊峻东等(2020)分析海州湾鹰爪虾(Trachysalambria curvirostris)资源开发率时发现E0.1与Gulland(1983)提出的最适开发率0.5更为接近; 刘逸文等(2020)采用不同方法来确定山东近海口虾蛄(Oratosquilla oratoria)资源开发率时, 发现E0.1变化相对较小, 是相对保守稳定的参考点。综上分析, 本研究采用E0.1作为海州湾毛虾资源开发率的目标参考点, 2020、2021年E0.1分别为0.60、0.77, 因此可通过调节渔业管理的两个重要管控要素(F、Lc)来达到渔业资源的最佳利用率, 以期获得最大可持续渔获产量(Ricker, 1975)。目前海州湾毛虾2020年和2021年开捕体长分别为26.28、29.73 mm, 计算所得出2020、2021年最适开捕体长Lopt分别为33.26、32.22 mm。如图 8所示, 当前海州湾毛虾开发状态处于A点, 且2020年和2021年开发率(E)分别为0.53、0.31, Lc/L∞分别为0.50、0.58, 对应的开捕体长(Lc)分别为26.28、29.73 mm。海州湾毛虾理论上的最佳开发状态为B点, 在M/K为1.83、1.79不变的情况下, 单位补充量渔获量Y′/R达到最大值0.035、0.045, 其对应的开发率E为0.60、0.77, Lc/L∞为0.62、0.63。若从A点调到B点, 单位补充量渔获量Y′/R将达到最大值, 其对应的开发率E增大13.2%、148%及Lc/L∞增大26.35%、9.34%, 而开发率的增加则对应着捕捞强度的增大。两种情况比较发现, 增大Lc对渔获量的影响较小, 而适当加大捕捞强度对渔获量提高影响较明显。2020年江苏连云港毛虾限额捕捞成功试点, 2021年山东省、辽宁省相继获批实施毛虾限额捕捞; 鉴于我国毛虾限额捕捞规模不断扩大, 应谨慎增加捕捞努力量, 将开发率有效控制E0.1以内, 做到“保护中利用、利用中保护”, 确保毛虾资源的可持续利用。
4 结论本研究首次探究了海州湾中国毛虾种群生物学特征及其资源开发状态, 研究发现相较于我国黄渤海海域, 6~7月繁殖期海州湾毛虾繁殖个体最大; 2020年和2021年海州湾毛虾资源均未过度开发。鉴于我国毛虾限额捕捞规模不断扩大, 应谨慎增加捕捞努力量, 将开发率有效控制在E0.1以内, 做到“保护中利用、利用中保护”。
参考文献
王毅波, 孙延瑜, 王彩霞, 等, 2019. 夏季渤海网采浮游植物群落和叶绿素a分布特征及其对渔业资源的影响. 渔业科学进展, 40(5): 42-51 |
仲崇峻, 曾晓起, 任一平, 等, 2001. 莱州湾、黄河口水域毛虾渔业生物学特征的研究. 海洋湖沼通报, (1): 31-36 |
伊峻东, 刘群, 2020. 海州湾及邻近海域鹰爪虾的生长、死亡参数及开发状况研究. 中国海洋大学学报, 50(12): 40-49 |
刘逸文, 张崇良, 刘淑德, 等, 2020. 山东近海口虾蛄单位补充量渔获量评估. 水产学报, 44(2): 213-221 |
刘瑞玉, 1956. 黄海和渤海的毛蝦(甲壳纲, 十足目, 樱蝦科). 动物学报, 8(1): 29-40 |
江苏省人民政府, 2020. 我省毛虾捕捞试点成效初显[EB/OL]. (2020-08-06)[2022-04-28]. http://www.jiangsu.gov.cn/art/2020/8/6/art_60085_9352202.html.
|
汤晓建, 宋大德, 吴磊, 等, 2019. 东海和黄海南部葛氏长臂虾资源状况及养殖潜力概述. 海洋渔业, 41(6): 744-752 |
李国东, 仲霞铭, 熊瑛, 等, 2021. 基于北斗船位数据的渔业信息解译与应用研究——以中国毛虾限额捕捞管理为例. 海洋与湖沼, 52(3): 746-753 |
李星颉, 戴健寿, 吴常文, 1986. 浙江北部沿岸海域的虾类资源. 浙江水产学院学报, 5(1): 13-20 |
杨晓改, 2015. 海州湾及其邻近海域浮游生物群落结构及其与环境因子的关系[D]. 青岛: 中国海洋大学.
|
宋海棠, 俞存根, 薛利建, 2012. 东海经济虾蟹类渔业生物学. 北京: 海洋出版社, 133-137
|
张孟海, 1986. 渤海湾、莱州湾毛虾生长速度和季节体长组成. 海洋湖沼通报, 4(13): 69-76 |
张孟海, 1992. 渤海湾、莱州湾毛虾的生殖特性. 海洋湖沼通报, (2): 58-67 |
张胜茂, 杨胜龙, 戴阳, 等, 2014. 北斗船位数据提取拖网捕捞努力量算法研究. 水产学报, 38(8): 1190-1199 |
陈立婧, 杨帆, 仲霞铭, 等, 2022. 中国毛虾生活史研究进展. 上海海洋大学学报, 31(5): 1032-1040 |
陈俅, 1981. 辽东湾中国毛虾资源的现状及渔业调整. 水产科学, (1): 13-17 |
罗文强, 赵刚, 张彦彦, 等, 2021. 海州湾海洋牧场人工鱼礁区建设前后海洋环境变化分析. 海洋湖沼通报, (1): 33-40 |
施仁德, 1986. 浙南近海中国毛虾Acetes chinensis Hansen的洄游分布及世代交替. 东海海洋, 4(1): 56-61 |
贺舟挺, 张亚洲, 徐开达, 等, 2012. 东、黄海黄(鮟)(鱇)的摄食习性及其随发育的变化. 渔业科学进展, 33(2): 9-18 |
徐从军, 徐宾铎, 张崇良, 等, 2019. 基于SURF指数识别海州湾食物网的关键饵料生物. 生态学报, 39(24): 9373-9378 |
徐从军, 隋昊志, 徐宾铎, 等, 2021. 基于LIM-MCMC模型研究海州湾食物网能量流动特征. 中国水产科学, 28(1): 66-78 |
徐君义, 1986. 乐清湾脊尾白虾的繁殖习性. 水产科技情报, (5): 22-24 |
殷名称, 1995. 鱼类生态学. 北京: 中国农业出版社, 57-61
|
郭斌, 张波, 金显仕, 2010. 黄海海州湾小黄鱼幼鱼的食性及其随体长的变化. 中国水产科学, 17(2): 289-297 |
韩耀全, 施军, 吴伟军, 等, 2017. 广西内陆水域主要增殖放流鱼类生长生物学特征及变化. 江苏农业科学, 45(16): 155-160 |
曾现英, 吴宁, 2016. 渤海湾、莱州湾毛虾季节分布与环境因子的关系. 河北渔业, (9): 15-16, 28 |
詹秉义, 1995. 渔业资源评估. 北京: 中国农业出版社, 1-353
|
BEVERTON R J H, HOLT S J, 1957. On the Dynamics of Exploited Fish Populations. London: Her Majesty's Stationery Office, 1-154
|
CLAUDIA C O, MIGUEL R S, MIGUEL ANGEL O N, et al, 2004. Effect of density and sex ratio on gonad development and spawning in the crayfish Procambarus llamasi. Aquaculture, 236(1/2/3/4): 331-339 |
GULLAND J A, 1983. Fish Stock Assessment: A Manual of Basic Methods[M]. Wiley: 223.
|
HOGGARTH D D, ABEYASEKERA S, ARTHUR R, et al, 2006. Stock assessment for fishery management: A framework guide to the use of the FMSP fish stock assessment tools[R]. Rome: FAO.
|
LIU D Y, WANG Y Q, 2013. Trends of satellite derived chlorophyll-a (1997-2011) in the Bohai and Yellow Seas, China: Effects of bathymetry on seasonal and inter-annual patterns. Progress in Oceanography, 116: 154-166 |
MAUCHLINE J, 1980. Measurement of body length of Euphausia superba Dana[C] //ANON. Biomass Handbook. Cambridge: Scientific Committee on Antarctic Research: 1-9.
|
OH C W, JEONG I J, 2002. Fisheries Biology of shrimps in the south western waters of Korea-1. Species composition of catches and spawning season of Acetes sp. for the Korean shrimp fishery. Korean Journal of Fisheries and Aquatic Sciences, 35(3): 223-230 |
OH C W, JEONG I J, 2003. Reproduction and population dynamics of Acetes chinensis (Decapoda: Sergestidae) on the western coast of Korea, Yellow Sea. Journal of Crustacean Biology, 23(4): 827-835 |
PAULY D, 1983. Length-converted catch curves: a powerful tool for fisheries research in the Tropics (part 2). Fishbyte, 2(1): 17-19 |
PAULY D, 1990. Length-converted catch curves and the seasonal growth of fishes. Fishbyte, 8(3): 33-38 |
RAHMAN M, OHTOMI J, 2018. Recruitment, growth patterns, and longevity of the deep-water velvet shrimp Metapenaeopsis sibogae (De Man. 1907) (Decapoda: Dendrobranchiata: Penaeidae). Journal of Crustacean Biology, 38(5): 552-562 |
RICKER W E, 1975. Computation and interpretation of biological statistics of fish populations[M]. Bulletin of the Fisheries Research Board of Canada: 209-210.
|
SONG X K, GU Y Z, ZHAI F G, et al, 2021. Climatology and seasonal variability of satellite-derived chlorophyll a around the Shandong Peninsula. Journal of Oceanology and Limnology, 39(4): 1222-1244 |
WEI Q S, WANG B D, FU M Z, et al, 2020. Spatiotemporal variability of physical-biogeochemical processes and intrinsic correlations in the semi-enclosed South Yellow Sea. Acta Oceanologica Sinica, 39(10): 11-26 |
WOOTTON R J, 1973. The effect of size of food ration on egg production in the female three-spined stickleback, Gasterosteus aculeatus L.. Journal of Fish Biology, 5(1): 89-96 |