 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 史兴宇, 王珍岩, 高微, 黄海军. 2023.
- SHI Xing-Yu, WANG Zhen-Yan, GAO Wei, HUANG Hai-Jun. 2023.
- 西太平洋卡罗琳M4海山区悬浮体分布及其影响因素
- DISTRIBUTION OF SUSPENDED PARTICULATE MATTER IN THE CAROLINE M4 SEAMOUNT IN THE WESTERN PACIFIC OCEAN AND ITS INFLUENCING FACTORS
- 海洋与湖沼, 54(3): 703-717
- Oceanologia et Limnologia Sinica, 54(3): 703-717.
- http://dx.doi.org/10.11693/hyhz20220800219
文章历史
-
收稿日期:2022-08-22
收修改稿日期:2022-10-20




2. 崂山实验室 海洋矿产资源评价与探测技术功能实验室 山东青岛 266237;
3. 中国科学院大学 北京 100049;
4. 中国科学院 海洋大科学研究中心 山东青岛 266071
2. Laboratory for Marine Mineral Resources, Laoshan Laboratory, Qingdao 266237, China;
3. University of Chinese Academy of Sciences, Beijing 100049, China;
4. Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
海洋悬浮体(suspended particulate matter, SPM)是自然海水中呈悬浮态存在的细小颗粒物的统称(Turner et al, 2002; 王珍岩等, 2017)。根据悬浮体的物质成分差异可以将其分为由浮游生物及其产生的有机包膜、粪球、部分生物碎屑等组成的有机颗粒和由矿物颗粒、无机生物骨骸等组成的无机颗粒(Bishop et al, 1980; 杨作升等, 1992)。悬浮体是自然海洋环境的重要组成部分, 也是海洋中物质和能量循环的重要载体, 在全球碳循环过程中发挥重要作用。海洋悬浮体的物质组成、分布和扩散输运过程与其所处的物理海洋环境关系密切(Bazzano et al, 2014; 高微等, 2016), 是开展现代海洋沉积过程以及海洋生态环境研究的重要研究对象(Li et al, 2020, 2021; Gao et al, 2021)。
海山(seamount)是指深海大洋中隆起高度超过1 000 m、顶部面积较小的海底地形(United States Board on Geographic Names, 1981; Rogers, 1994)。海山与稳定洋流的相互作用可能会形成上升流、泰勒帽(Taylor cap)等特殊的物理过程, 在海山顶部形成等温线上凸的现象(Hogg, 1973); 海山与周期性潮流相互作用也会产生潮余流, 较强的潮余流能够在海山顶部引发反气旋环流, 形成与泰勒帽类似的等温线抬升现象(White et al, 2007)。西太平洋是全球海山最集中分布的海区之一(Yesson et al, 2021)。西太平洋海洋环流结构复杂, 主要由赤道流系、西边界流系和多条贯穿流等构成(Hu et al, 2015)。此外, 该海区还是多个来自高纬度、不同层位水团交汇的交叉路口(Kaneko et al, 2001)。洋流与海山相互作用改变了局地海洋动力环境, 不可避免地会对海山区海水温盐结构、营养盐分布、生态环境和沉积动力环境等产生巨大影响, 进而改变海洋物质和能量循环过程, 对维持大洋海区生物多样性、提升海洋碳循环能力等产生重要影响。因此, 开展西太平洋海山区悬浮体分布及其控制机制研究对于深入理解海山在大洋海区物质和能量循环过程中的角色和作用具有重要意义。
本文以西太平洋卡罗琳洋脊M4海山为研究对象, 对2017年夏季在该海山区开展水文环境和悬浮体调查获得的数据进行分析, 探讨地形作用、海水层化以及洋流输运等因素对海山区悬浮体分布的影响机制, 可为深入开展海山区沉积动力学研究和生态系统动力学研究提供参考和借鉴。
1 研究区域背景卡罗琳M4海山位于环流结构复杂的西太平洋海区。向西流动的北赤道流(north equatorial current, NEC)和它的两个分支——向北流动的黑潮(Kuroshio)和向南流动的棉兰老流(Mindanao current, MC)是赤道以北西太平洋表层流的重要组成部分, 而这三个表层流下方分别对应了向东流动的北赤道潜流(north equatorial undercurrent, NEUC; > 300 m)、向南流动的吕宋潜流(Luzon undercurrent, LUC; > 500 m)和向北流动的棉兰老潜流(Mindanao undercurrent, MUC; > 600 m) (Zhang et al, 2014, 2017; Wang et al, 2015) (图 1a)。
 |
| 图 1 西太平洋上层环流结构示意图(a)(Wang et al, 2015)及M4海山站位分布图(b) Fig. 1 Schematic of upper circulation structure in the tropical Western Pacific Ocean (a) and distribution of stations on the M4 seamount (b) 注: NEC: 北赤道流; Kuroshio: 黑潮; MC: 棉兰老流; NECC: 北赤道逆流; NGCC: 新几内亚沿岸流; ME: 棉兰老涡; HE: 哈马黑拉涡; NGCUC: 新几内亚沿岸潜流; MUC: 棉兰老潜流; LUC: 吕宋潜流; NEUC: 北赤道潜流; b中黑色虚线为水深等值线(单位: m); 红圈标识站位: 对悬浮体样品进行了扫描电镜及能谱分析 |
M4海山位于西太平洋卡罗琳岛脊北端, 该海山从2 800 m水深处隆起, 向上延伸到距离海洋表面50 m左右的真光层内部, 整体呈北北西走向。M4海山山顶面积较大, 最大直径可达约11 km, 整体呈现出边缘高、中心低的特征。海山坡度从坡底的10°左右缓慢增加到坡顶的大于40°, 且在山顶边缘突然增加到55°甚至超过60° (甘雨, 2021)。
2 材料和方法 2.1 现场观测与取样2017年8月15~26日, “科学号”海洋科考船在西太平洋卡罗琳岛脊的M4海山区(10.2°~10.8°N, 139.8°~140.4°E)开展了综合调查, 对该海区2个交叉断面(断面A和断面B, 交叉中心为A-0站)、共21个站位进行了观测和采样, 断面及站位的分布情况如图 1b。利用美国Sea-Bird公司生产的SBE 9 plus型温盐深传感器(conductivity-temperature-depth, CTD)和SBE 11 plus型甲板单元对各站位的水体温度(℃)、电导率(S/m)及压力(Pa)进行剖面观测。利用CTD外置的SBE 43型溶解氧传感器及WET Labs ECO-FL荧光叶绿素传感器获取了各站位水体的溶解氧浓度(mg/L)及荧光叶绿素浓度(mg/m3)。使用SBE Data Processing软件对上述获取的原始数据进行处理, 最后导出1 m平均的数据。美国TRDI公司生产的OS-38相控阵海洋调查型声学多普勒流速剖面仪(acoustic doppler current profiler, ADCP)被安装在船体上以记录1 000 m以浅的洋流。由于船体的影响, M4海山区的第一个有效海流数据位于55.07 m, 最大探测深度约1 000 m, 相邻数据间的深度间隔为32 m。利用WinADCP软件对原始数据进行质量控制并导出1 min平均的流速流向数据。
除了仪器测量数据外, 我们还采集了各站位代表性水层的海水样品进行现场过滤。过滤使用的滤膜为直径47 mm、孔径0.45 μm的混合纤维素酯滤膜, 每个样品过滤的海水体积在5~10 L之间, 过滤后用适量的蒸馏水滤洗以去除滤膜上的盐分。
2.2 实验室分析将悬浮体滤膜样品带回实验室后依次烘干、称重, 根据过滤水量及过滤前后滤膜质量的差异计算悬浮体质量浓度。然后选取部分代表性的样品(表 1), 利用扫描电镜(scanning electron microscope, SEM)与能谱仪(energy dispersive spectroscopy, EDS)进一步分析其物质组成。SEM-EDS为捷克TESCAN公司VEGA3型钨灯丝扫描电子显微镜, 配置德国Bruker Nano GmbH 610M型能谱仪。样品的准备及实验步骤如下:
| 站位 | 经度 | 纬度 | 水深/m |
| A-0 | 140.134 7°E | 10.477 0°N | 3, 15, 50, 101 |
| A-4 | 140.189 9°E | 10.304 1°N | 3, 50, 120, 150, 300, 500, 1 000, 1 506 |
| A-8 | 140.121 6°E | 10.663 0°N | 3, 50, 100, 128, 300, 500, 1 000, 2 000, 3 490 |
| B-5 | 139.927 9°E | 10.466 6°N | 3, 50, 100, 150, 300, 500, 1 000, 2 000, 2 763 |
| B-11 | 140.377 3°E | 10.532 1°N | 3, 50, 100, 131, 300, 500, 1 000, 2 000, 2 448 |
(1) 将待观察的滤膜样品放入烘箱内进行4 h的低温烘干, 然后将每张滤膜裁剪下一个小的扇形部分, 用双面碳导电胶带黏贴在易导电的金属样品台上, 在高真空条件下用喷金仪将样品喷金(Au)后放入扫描电子显微镜的样品舱内, 设置低真空模式, 调整工作距离为15 mm, 加速电压20.0 kV。
(2) 由于大洋悬浮体浓度较低, 在滤膜上基本不会重叠, 所以在每个样品上选取5个大小相同的视域(231 μm × 173 μm)逐一进行观察。视域的选择如图 2所示: 在靠近扇形样品圆心角的一侧选择第一个观察点, 然后沿扇形半径方向以约3 mm为间隔向外移动, 直到观察5个视域。
 |
| 图 2 样品中5个观察视域分布示意图 Fig. 2 Distribution of 5 views on a sample |
(3) 在每一个观察点先用背散射电子探测器获取该视域的背散射电子像, 在固定电镜的观察倍数的条件下, 转换到二次电子探测器获取同一视域下的普通二次电子像(图 3), 然后放大该视域中的小颗粒, 获取其图像同时可以进行能谱测试。
 |
| 图 3 同一视域下的背散射电子像(a)及二次电子像(b)示例 Fig. 3 Examples of backscattered electron image (a) and secondary electron image (b) in a same view |
实验结束后, 我们根据获取的图像及能谱分析数据鉴别滤膜上悬浮体的种类并对每种类型悬浮体进行计数, 对每个站位不同水层的5幅图像中的悬浮体数量取均值最后结合悬浮体样品的观察面积、过滤面积以及实际滤水量计算出每种类型悬浮体的丰度(单位: 个/L)。
3 结果 3.1 温盐及荧光叶绿素a浓度分布从图 4e可以看出, 调查期间M4海山区的表层流及其下方的海流存在明显的差异, 通过Shi等(2021)对该海山区海流的详细分析可知, M4海山区200 m以浅为自东向西流动的北赤道流, 流速较高, 范围在0.10~0.35 m/s之间; 而300 m以深大部分站位的海流均为自西向东流动, 流速相对200 m以浅略有降低, 范围在0.05~0.20 m/s之间, 明显受到了北赤道潜流的控制。
 |
| 图 4 M4海山区2个断面0~150 m (a, b)及0~1 000 m (c, d)的位势密度(黑色曲线, kg/m3)和温度分布以及海山区各站位的平均海流剖面(e) Fig. 4 Distributions of potential density (black curves, kg/m3) and in-situ temperature between 0~150 m (a, b) and 0~1 000 m (c, d), and the mean current profile (e) of the M4 seamount |
从图 4和图 5可以明显看出, M4海山区的温度和盐度在垂向上存在明显分层现象。2017年夏季, M4海山区海表面温度较高, 最高可达30.87 ℃, 北赤道流影响范围内(0~200 m)海水温度随水深的增大而迅速降低, 200 m处水体的温度约为15 ℃; 温度跃层位于60~180 m之间(图 4c~4d)。0~200 m水深之间的盐度呈现出先升高后降低的趋势, 海表面盐度仅有约33.50, 在水深100~200 m之间存在一个盐度高值区, 最高盐度可达35.11, 这一高盐水体为北太平洋热带水(north Pacific tropical water, NPTW)(Shi et al, 2021); 表层水体与北太平洋热带水的盐度差异使得水深30~ 200 m之间形成了盐度跃层, 其中100 m水深附近由于北太平洋热带水的平流和延伸作用形成了强烈的盐度梯度, 因此该位置的盐度跃层强度最大(图 5c~5d)。
 |
| 图 5 M4海山区2个断面0~150 m (a, b)、0~300 m (c, d)及300~1 000 m (e, f)的盐度分布 Fig. 5 Distributions of salinity in 0~150 m (a, b), 0~300 m (c, d), and 300~1000 m (e, f) of the M4 seamount |
在北赤道潜流流经的范围内(300~800 m), 水体温度随着水深的增大缓慢降低, 800 m以深的温度在5 ℃左右(图 4c~4d); 该范围内的盐度在北太平洋中层水和南极中层水共同的影响下(Shi et al, 2021)呈现出在海山各个方向上不均匀的分布特征(图 5e~5f)。
M4海山区表层水体的荧光叶绿素a浓度较低, 所有站位的平均值为0.100 mg/m3。0~200 m之间的荧光叶绿素a浓度先升高后降低, 在80~170 m之间形成次表层叶绿素最大值层(subsurface chlorophyll maximum layer, SCML), 浓度值最大为0.910 mg/m3, 各站位SCML的深度(荧光叶绿素a浓度最大值所在的深度)位于107~141 m之间(水深小于100 m的A-0、A-1及A-5站除外)。200~1 000 m之间的荧光叶绿素a浓度均低于0.200 mg/m3, 且其随水深的变化很小(图 6c~6d)。这一结果与Dai等(2020)利用10-AU Turner Design荧光计测量的2017年夏季M4海山区叶绿素a浓度的分布趋势一致。
 |
| 图 6 M4海山区2个断面0~150 m (a, b)及0~1 000 m (c, d)的荧光叶绿素a浓度分布 Fig. 6 Distributions of fluorescence chlorophyll a in 0~150 m (a, b) and 0~1000 m (c, d) of the M4 seamount |
除了上述特征之外, 调查期间M4海山山顶上方还存在明显的等值线抬升现象。M4海山山顶大致位于50~100 m之间, 因此海山对这一范围内的水体影响显著。图 4a显示, M4海山A断面上山顶的位势密度等值线明显抬升, 在山顶上方形成冷穹, B断面山顶上方的等位势密度线也存在明显的隆起和下凹(图 4b); 2个断面上山顶附近的盐度也表现出与位势密度及温度相似的分布特征(图 5a~5b)。Shi等(2021)通过理论计算和分析认为M4海山山顶上方位势密度、温度及盐度的抬升是由于潮流和海山的相互作用产生的。除了位势密度、温度和盐度之外, 我们也观察到在该海山区的50~75 m水深之间, A断面的A-5站位和B断面的B-2、B-7、B-8和B-9站位的荧光叶绿素a浓度明显高于其他站位(图 6a~6b), 因此山顶冷穹的形成很可能对这一水层中浮游生物的分布产生了影响。
3.2 悬浮体物质组成及分布特征M4海山区各站位的悬浮体质量浓度剖面(图 7)显示, 水体中的悬浮体质量浓度整体偏低, 范围在0.00~0.70 mg/L之间。在垂向上, 各个站位悬浮体质量浓度总体表现出随水深先增大后减小的分布特征, 浓度高值大多出现在200 m水深以浅的范围内。
 |
| 图 7 M4海山区各站位悬浮体含量、温度及盐度分布 Fig. 7 Distributions of SPMs content, temperature, and salinity at each station of the M4 seamount |
扫描电镜及能谱分析结果表明, M4海山区水体中的悬浮体可以分为生物碎屑、矿物颗粒和絮凝体等3类。
3.2.1 生物碎屑在本研究中我们将所有单独分布于滤膜上的、具有明显生物特征颗粒均归为生物碎屑, 而不考虑这些颗粒的磨损及破碎程度。M4海山区的悬浮体样品中分布有较多的生物碎屑, 它们大小不一, 种类各异。较小的生物碎屑粒径小于10 μm, 较大的可达几百微米, 多数生物碎屑颗粒的粒径大小在5~30 μm之间。由于滤膜样品经过了抽滤、烘干, 能够保留在滤膜上的生物大多数为具有坚固的硅质或钙质细胞壁的生物, 因此滤膜上可见的生物碎屑以硅藻为主(图 8a, 8b), 占总生物碎屑的90%, 除硅藻外, 甲藻(图 8c)及颗石藻(图 8d)比较常见, 它们均占生物碎屑总数的4%, 剩余的生物碎屑包括放射虫、硅鞭藻等仅占生物碎屑总数的2%。
 |
| 图 8 M4海山区水体中的典型悬浮体 Fig. 8 Typical SPMs in water of the M4 seamount 注: a. 菱形藻; b. 圆筛藻; c. 甲藻; d. 颗石藻; e. 硅酸盐矿物; f. 碳酸盐矿物; g. 氧化物; h. 自生矿物; i. 有机包膜; j. 矿物絮凝体; k. 生物絮凝体; l. 混杂絮凝体 |
M4海山区5个站位不同水层的生物碎屑丰度在5.55×103~5.82×104个/L之间。离山顶较远站位(除海山西侧的B-5站外)的生物碎屑丰度随水深的增大先升高后降低, 峰值位于SCML中, 但各站位的丰度最大值相差较大, 按生物碎屑丰度排序为A-4 > A-8 > B-11 > B-5; 山顶上方A-0站位生物碎屑丰度的垂向分布与其他站位存在明显差异, 该站位生物碎屑丰度的高值出现在表层及15 m水层, 最大值为5.82×104个/L, 是其他站位表层水体中生物碎屑丰度的3倍左右。300 m以深的水体中各站位的生物碎屑丰度随水深的增大波动变化, 其中海山西侧B-5站300 m以深的生物碎屑丰度最高(图 9)。
 |
| 图 9 M4海山区5个站位0~300 m以及0~3 500 m水体中的总悬浮体、生物碎屑、矿物颗粒以及絮凝体丰度的垂向分布 Fig. 9 Distributions in abundance of the total SPMs, organic debris, mineral particles, and flocs in 0~300 m and 0~3 500 m at 5 different stations of the M4 seamount |
本研究对矿物颗粒的识别和计数主要依赖背散射电子像及能谱分析。背散射电子产率随元素原子序数的增加而增加, 因此背散射电子像中矿物颗粒的亮度一般高于其他颗粒, 表现出更好的成分衬度。但由于背散射电子离开样品表面后沿直线运动, 背向探测器出射的背散射电子达不到探测器, 导致图像有阴影; 而背向探测器出射的二次电子可以通过弯曲的轨迹进入探测器, 虽然其信号会有所减弱, 但也有利于显示背向探测器部位的细节, 而不会形成阴影。因此二次电子像的形态衬度要优于背散射电子像, 能够显示更清晰的颗粒样貌。我们将背散射电子像中亮度明显较高的单体颗粒(除硅质及钙质生物外)均计为矿物颗粒。但由于大洋悬浮体中的矿物颗粒粒径较小, 能谱仪的扫描尺寸大于矿物颗粒的粒径, 导致扫描结果受到目标颗粒周围的其他颗粒、滤膜以及滤膜孔隙中小颗粒的影响, 难以准确地给出矿物的名称。因此只将样品中的矿物颗粒分为硅酸盐矿物、碳酸盐矿物、氧化物和氢氧化物以及一些特殊的自生矿物(图 8e~8h)。统计结果表明, M4海山区水体中的矿物颗粒的粒径多在1~10 μm之间, 且以硅酸盐矿物为主。
M4海山区水体中矿物颗粒的丰度远高于生物碎屑丰度, 5个站位不同水层中矿物颗粒丰度的范围在3.78×104~2.52×105个/L之间。除A-8站外, 其他4个站位3~100 m水深之间的矿物颗粒丰度基本随水深的增大而缓慢增大, 在100 m水深处形成一个小的峰值; A-0站位100 m以浅水体中的矿物颗粒丰度明显高于其他站位。150 m水层中的矿物颗粒丰度与100 m相比有所减小, 而150 m以深的水体中, 矿物颗粒丰度又随水深的增大而迅速增大, 在300~500 m水深之间的矿物颗粒丰度是表层及深层水体中(> 1 000 m)的2倍左右, 形成了一个明显的峰值; 500~1 000 m水体中的矿物颗粒丰度随水深的增大而减小; 1 000 m以深水体中矿物颗粒的丰度基本保持不变(图 9)。
3.2.3 絮凝体本研究中的絮凝体包含粪团、有机包膜、生物絮凝体、矿物絮凝体及混杂絮凝体, 但由于大部分有机包膜黏附在滤膜上无法分辨, 因此只对边缘清晰的有机包膜进行了计数。由于过滤过程的影响, 悬浮体滤膜上的絮凝体以有机包膜和粪团为主, 它们的粒径一般在20 μm以上(图 8i), 而生物絮凝体、矿物絮凝体及混杂絮凝体较少, 其粒径多在5~50 μm之间(图 8j~8l)。
M4海山区不同水层中絮凝体丰度的范围在2.70×104~2.19×105个/L之间。表层水体中的絮凝体丰度较高, 5个站位3 m层的平均丰度值可达9.15× 104±2.53×104个/L; 50 m层的絮凝体丰度有所减小, 平均值为5.90×104±1.86×104个/L; SCML中的絮凝体丰度有所增大, 特别是山顶上方A-0站位101 m水层的丰度达到2.19×105个/L; 150~300 m水体中的絮凝体丰度随水深的增大缓慢增大; 而300 m以深各站位的絮凝体丰度整体上变化不大(图 9)。
前人研究表明, 生物作用是影响大洋悬浮体垂向结构的重要因素, 浮游生物及其代谢产物等是表层悬浮体的主要物质来源(Cho et al, 1990; 杨作升等, 2007; Turner, 2015)。M4海山区水体的荧光叶绿素a浓度高值主要位于200 m以浅(图 6), 与悬浮体质量浓度的高值分布范围一致(图 7), 二者均呈现出随水深先增大后减小的分布趋势, 因此, 浮游植物是造成M4海山区表层悬浮体质量浓度高值的主要原因。从颗粒组分的角度, 悬浮体质量浓度的垂向分布特征与生物碎屑较为一致, 而与矿物颗粒相差较大(图 9)。这是由于该海山区的生物碎屑主要由密度较大的硅藻组成(90%), 尽管其丰度较低, 但硅藻的粒径普遍大于矿物颗粒, 此外, 絮凝体主要由密度较小的有机包膜组成。因此, 尽管统计结果表明矿物颗粒及絮凝体的丰度占总悬浮体丰度的比重较大, 但它们对悬浮体质量浓度的贡献远小于生物碎屑。
4 讨论 4.1 海水层化对悬浮体组成和分布的影响前人研究表明, 西太平洋暖池海表面温度高于28 ℃, 盐度较低, 硝酸盐和叶绿素浓度非常低, 表现为明显的寡营养特征(Le Borgne et al, 2002; Messié et al, 2006)。在寡营养海域, 上层水体中的营养盐浓度全年处于较低水平, 营养盐结构极度不平衡, 限制了浮游植物的生长, 从而影响了表层悬浮体的物质组成, 同时降低了表层悬浮体的丰度。一般认为, 造成暖池区寡营养条件的因素主要有两个: 一是上层水体中的浮游植物大量消耗营养物质, 导致营养盐浓度急剧降低; 二是海水的强烈层化限制了含有高浓度营养盐水体的向上输送, 使得跃层以浅的水体营养盐浓度极低(Murray et al, 1994; Ma et al, 2021b)。
Ma等(2021b)研究了小型浮游生物对M4海山周围海域寡营养条件的影响。他们通过分析原绿球藻、聚球藻和细菌的丰度在垂向上的分布与营养盐之间的关系, 发现聚球藻和细菌的丰度与营养盐浓度呈明显的负相关关系。聚球藻和细菌都具有较强的环境适应性, 能够在寡营养环境中吸收营养物质, 聚球菌还能够为细菌的生长提供充足的碳源。因此Ma等(2021b)认为聚球藻和细菌共同加剧了M4海山区上层水体营养物质的缺乏, 是导致该海域寡营养特征的重要因素之一。
除了浮游生物的影响, 海水的强烈层化也可以阻碍上、下层水体之间营养物质的交换, 使上层水体表现为寡营养特征。从前文的分析可知, 现场调查期间M4海山区的水体分层明显, 水深30~200 m之间分布着盐度跃层, 60~180 m之间分布着温度跃层。温度跃层和强盐度跃层的共同作用, 阻碍了水体的垂直交换, 导致M4海山区上层水体中的营养盐浓度极低。从M4海山区的营养盐垂向分布特征显示, 营养盐浓度从100 m水深开始迅速增加(Ma et al, 2019), 这一位置与北太平洋热带水的起始水深一致, 说明跃层的强度越大, 对营养盐的阻碍作用越明显。
海水的强烈层化使得M4海山区深水中的高浓度营养盐难以向上扩散, 导致上层水体中的营养盐浓度极低, 进而限制了表层水体中浮游植物的生长, 因此浮游植物生物量很低(Dai et al, 2020), 荧光叶绿素a浓度也处于较低水平(< 0.2 mg/m3, 图 6)。此外, M4海山区水深3~100 m之间的悬浮体(特别是矿物颗粒)丰度随水深的增加而缓慢增大, 并且在100 m水深处形成了一个较小的峰值(图 9)。这是由于表层的悬浮体在沉降过程中受到了温度跃层和盐度跃层的双重阻碍, 随着水深的增大, 跃层强度增大, 而悬浮体的下沉速率逐渐降低, 最终悬浮体在跃层强度最大的水层(100 m水深附近)逐渐积累, 从而出现悬浮体丰度的峰值。
随着深度的增大, 跃层强度减弱, 营养盐浓度逐渐升高(Ma et al, 2019), 形成了有利于浮游植物生长的条件, 因此100 m以深的浮游植物生物量与表层相比明显增加(Dai et al, 2020), 荧光叶绿素a浓度也明显升高, 形成了SCML。由于浮游生物生物量增加, 生物分泌的有机质也增多, 因此该水层中的生物碎屑和絮凝体也表现出较高的丰度(图 9)。在SCML以深的水体中, 尽管营养盐浓度依旧随水深的增大不断升高, 但氮与磷的比值明显降低, 形成了氮限制(马骏, 2020), 再加上光照强度的减弱, 限制了浮游植物的生长, 使得荧光叶绿素a浓度明显降低。这一水深的悬浮体丰度, 特别是生物碎屑和絮凝体的丰度也明显降低(图 9, 150 m)。M4海山区悬浮体质量浓度的高值主要位于200 m以浅的水体中, 并且在垂向上表现出随水深先增大后减小的分布趋势(图 7), 与上述浮游植物的分布一致, 说明温盐跃层通过控制营养盐的垂向分布影响了浮游植物的生长, 而浮游植物产生的生物碎屑是200 m以浅水体中悬浮体质量浓度的主要组成部分, 因此这一水层中悬浮体质量浓度的分布特征与浮游植物一致。
4.2 海山地形对悬浮体分布的影响海山地形与洋流的相互作用能够产生一些独特的物理过程, 进而对海山的生态系统产生影响(White et al, 2007; Lavelle et al, 2010)。2017年夏季, M4海山山顶的温度及盐度等值线向上抬升, 形成了一个明显的冷穹。前人研究认为, 海山山顶的冷穹可能是由泰勒帽或潮调整(tidal rectification)产生的。Shi等(2021)通过理论计算认为M4海山区不具备形成泰勒帽的条件, 因此调查期间山顶的冷穹不是由泰勒帽产生的; 同时, 由于山顶附近的温盐结构与潮调整产生的环流结构一致性很高, 因此Shi等(2021)认为潮调整是M4海山山顶产生冷穹的主要机制。
为了进一步确认潮流与海山的相互作用在冷穹形成中的贡献, 我们通过公式(1) (Wright et al, 1985)计算潮调整过程中产生的最大潮余流:
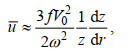 (1)
(1)式中, V0是潮汐强迫的振幅(此处采用M2分潮的平均最大流速), ω为潮汐强迫的角频率, f为局部科氏参数, z为局部水深, 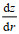
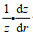
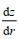
在其他海山区的研究中也观察到了山顶冷穹或反气旋环流的存在。Genin等(1985)在西北太平洋的Minami-Kasuga海山(21°36′N, 143°38′E; 260 m)山顶发现等温线向上抬升到较浅的透光层; Dower等(1992)在东北太平洋的Cobb海山(46°46′N, 130°48′W; 24 m)观察到了反气旋环流的存在; Read等(2017)在西南印度洋深度较浅的Melville Bank和Coral海山顶部都发现了明显的等温线及等密度线的抬升。而反气旋环流和冷穹的存在, 也引起了海山生态系统的改变。Minami-Kasuga海山的冷穹内不仅出现了相对较高的叶绿素浓度, 冷穹内和冷穹上方的浮游动物生物量也更高(Genin et al, 1985); Cobb海山的反气旋环流内(山顶周围)也观察到了叶绿素水平的提高(Dower et al, 1992); Melville Bank和Coral海山山顶冷穹内的POC浓度低于周围海域, 可能是较强的生物活动造成的(Djurhuus et al, 2017)。这些研究都证明了海山山顶的冷穹(反气旋环流)能够对其周围的生物活动和颗粒物的分布等产生明显的影响。
由前文的分析可知, 在M4海山区的50~75 m水深之间, A-5、B-2、B-7、B-8和B-9站位(冷穹内)的荧光叶绿素a浓度明显高于冷穹外的站位(图 6a, 6b)。荧光叶绿素a浓度的高低往往能够体现一个海区的浮游生物量水平(Martin et al, 2010; Brotas et al, 2013), 荧光叶绿素a浓度的升高说明冷穹内悬浮体中的浮游生物组分也有所增加。这一现象证明, 在反气旋环流的作用下, 营养盐浓度较高的深层水体突破了较强的跃层, 扩散到寡营养的上层水体中, 促进了海山顶部浮游生物的生长, 因此在M4海山区50~75 m的水层中, 冷穹内的荧光叶绿素a浓度高于远离海山的站位。
除了荧光叶绿素a浓度之外, M4海山区不同站位的悬浮体丰度也存在明显的差异。与远离山顶(冷穹外)的站位相比, M4海山山顶A-0站位(冷穹内)的总悬浮体丰度和4种不同类型的悬浮体丰度均显著增大(图 9~10)。这些现象都说明在M4海山区, 潮调整产生的反气旋环流对悬浮体起到了明显的滞留作用。M4海山区200 m以浅为自东向西流动的北赤道流, 因此山顶位置增加的悬浮体部分来源于北赤道流的水平输运; 反气旋环流还能够将山顶下方SCML中浓度较高的悬浮体向上输运, 这部分颗粒也会被冷穹滞留在海山山顶。因此, A-0站的悬浮体丰度远高于离海山较远的其他站位。
 |
| 图 10 M4海山区3, 50及100 m水层中总悬浮体丰度之和(S)以及3种不同类型悬浮体丰度各自的和在不同站位的分布 Fig. 10 The sum (S) of total abundance of SPMs at 3 m, 50 m, and 100 m water layers of the M4 seamount and the distribution of the respective sum of four different types of suspended particle abundance in different stations 注: 等值线表示水深(单位: m) |
M4海山山顶反气旋环流的滞留作用不仅表现在荧光叶绿素a浓度和悬浮体丰度的分布上。多学科研究表明, 2017年夏季M4海山区山顶站位的NO3-N浓度、PO4-P浓度、细菌丰度、分级叶绿素a浓度以及浮游病毒丰度等都明显高于离山顶较远的站位(Dai et al, 2020; Zhao et al, 2020; Ma et al, 2021a)。这些现象进一步证明了M4海山山顶存在反气旋环流, 它对周围海域的营养盐、悬浮体和浮游生物等都产生了明显的滞留效应。
4.3 洋流对陆源颗粒的输运M4海山区5个站位不同类型悬浮体丰度的垂向分布显示, 该海山区各站位水体中的总悬浮体丰度、矿物颗粒丰度和絮凝体丰度的极大值均出现在300~ 500 m之间的水层中(图 9), 并且这一深度与北赤道潜流的核心位置非常一致(图 4e)。
从前文的分析可知, 由于M4海山区上层海水层化强烈, 从海洋表面输入的风尘颗粒向下沉降时受到了温度跃层和盐度跃层的阻碍, 下沉速率逐渐降低, 并在100 m水深附近逐渐积累而形成了一个小的丰度峰值(图 9), 这部分矿物颗粒很难继续向下沉降。同时前人的研究指出, 风尘而来的陆源颗粒在深层水体中的分布一般较为均匀(高微, 2018), 而M4海山区300~500 m之间的矿物颗粒丰度远高于100 m水层的矿物颗粒丰度, 因此M4海山区300~500 m水深之间高丰度的矿物颗粒不是从浅水沉降而来的。
前人研究发现, 复杂的西边界流系可以通过输送陆源物质影响西太平洋海区海水的营养盐水平, 对该海区的物质循环产生重要的影响(Lukas et al, 1996; Kashino et al, 2015), 而西边界流系对陆源颗粒的输运过程尚未引起足够的关注。高微(2018)对2014年冬季在西太平洋海区获取的悬浮体样品进行了同步辐射微束X射线荧光分析(synchrotron radiation X-ray fluorescence, SRXRF), 得到了悬浮体样品的元素分析数据, 发现位于北赤道潜流流经位置的I-8站位(图 1a) 300 m水层中悬浮体陆源元素(Ti, Mn, Fe, Zr)的含量极高, 同时该深度恰好与棉兰老潜流挟带来的中层高盐水的位置一致(图 11)。她通过详细分析西边界流系的流动路径及其所挟带的水团特性, 认为是棉兰老潜流和北赤道潜流共同将大量的陆源物质输送到了I-8站位, 因此该站位300 m水层悬浮体的陆源元素含量极高。除了I-8站位之外, 与其同一时间调查的雅浦Y3海山(8.7°~9.1°N, 137.6°~137.9°E; 图 1a) 300 m水层中悬浮体的Fe、Mn元素含量也极高(图 11), 且Y3海山区的300~700 m之间也发现了高盐的水团(Wang et al, 2022)。这些现象说明太平洋西边界的陆源物质很可能通过西边界流系到达距离陆地较远的大洋海域。

由前人的研究可知, 太平洋西边界的新几内亚沿岸潜流(New Guinea coastal undercurrent, NGCUC)向下延伸到南极中层水所在深度, 之后挟带南极中层水跨过赤道(Tsuchiya, 1991), 汇入棉兰老潜流(Qu et al, 2004), 王凡等(1997)发现棉兰老潜流总流量的60%左右为南极中层水。棉兰老潜流向北扩散, 并在9.5°~10.5°N之间分为两部分, 一部分继续向北流动, 另一部分则向东汇入北赤道潜流(Wang et al, 2015)。在新几内亚沿岸潜流和棉兰老潜流的流动路径上, 包括新几内亚岛以及菲律宾群岛在内的地区, 每年的泥沙入海量都很大(Milliman et al, 2013)。因此, 新几内亚沿岸潜流和棉兰老潜流能够挟带大量的陆源物质向北流动, 然后向东汇入北赤道潜流。由此可知, 在西边界流系和赤道流系的输运下, 菲律宾沿岸甚至南太平洋的岛屿都能够为西太平洋海域供给陆源物质。
M4海山区位于9.5°~10.5°N之间的北赤道潜流的流动路径上, 由图 12a可知, 该海山区300~700 m之间水体的T-S特征不同于典型的北太平洋中层水, 而是表现出稀释的南极中层水的T-S特征; 同时, 这一水层的溶解氧浓度略高于其上方和下方的水体(图 12b)。以上现象都证实, 由棉兰老潜流挟带的高盐、高溶解氧的南极中层水部分转向东, 并通过北赤道潜流进入了M4海山所在海域。而在M4海山区, 北赤道潜流的核心位置(流速最大的深度)在300~600 m水深之间(图 4e), 与该海区矿物颗粒丰度峰值所处的水层一致。因此, M4海山区300~500 m之间高丰度的悬浮矿物颗粒很可能是由棉兰老潜流和北赤道潜流输运来的。
 |
| 图 12 M4海山区中层水体的温盐散点图(a)及5个站位0~1 000 m之间的溶解氧浓度剖面(b) Fig. 12 The T-S scatter diagram of intermediate water of the M4 seamount (a) and the profile of dissolved oxygen concentration in 0~1 000 m at the 5 stations 注: a中虚线为位势密度等值线(单位: kg/m3), 2条黑色粗线分别代表北太平洋中层水(左)和南极中层水(右)的典型T-S关系曲线 |
我们推测: 太平洋西边界的新几内亚沿岸潜流挟带了大量来自新几内亚岛的陆源物质向北流动并汇入棉兰老潜流, 棉兰老潜流在向北流动的过程中也带走了菲律宾群岛沿岸的陆源颗粒, 在9.5°~10.5°N之间, 棉兰老潜流产生2个不同的分支, 向东的一支挟带着部分陆源颗粒汇入北赤道潜流, 而北赤道潜流又挟带着这些陆源颗粒到达M4海山区, 使得该海山区300~500 m水体中的矿物颗粒丰度显著增加。这一推测还需要进一步的调查数据来证实。
5 结论本文对2017年夏季在西太平洋卡罗琳M4海山区获得的水文及悬浮体资料进行分析, 得到如下结论:
(1) M4海山区海水的强烈层化使得深水中的高浓度营养盐难以向上扩散, 导致上层水体中的营养盐浓度极低, 限制了浮游植物的生长, 使得荧光叶绿素a浓度处于较低水平, 跃层的存在也阻碍了悬浮体下沉, 使得悬浮体在跃层上方逐渐积累; 跃层下方营养盐浓度逐渐升高, 形成了有利于浮游植物生长的条件, 荧光叶绿素a浓度明显升高, 出现了SCML, 浮游生物的增加使SCML悬浮体中的生物碎屑和絮凝体丰度也升高。
(2) 潮调整在M4海山顶部形成了反气旋环流, 在反气旋环流的作用下, 营养盐浓度较高的深层水体突破了跃层, 扩散到寡营养的上层水体中, 促进了海山顶部浮游生物的生长, 使得荧光叶绿素a浓度升高; 反气旋环流也对不同来源的悬浮体起到了明显的滞留作用, 导致山顶站位100 m以浅水体中的悬浮体丰度远高于离山顶较远的站位。
(3) 在新几内亚沿岸潜流和棉兰老潜流的输运下, 菲律宾群岛沿岸的陆源矿物颗粒向北扩散, 并向东汇入北赤道潜流, 最终到达M4海山区, 使得该海山区300~500 m水体中的矿物颗粒丰度显著增加。
马骏, 2020. 西太平洋海山区"海山效应"对海水生源要素影响的探析[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 40-42.
|
王凡, 胡敦欣, 1997. 棉兰老潜流水源的初步研究[M] //叶笃正. 赵九章纪念文集. 北京: 科学出版社: 345-352.
|
王珍岩, 张洪格, 高微, 2017. 冬季西太平洋雅浦海山(Y3)区次表层叶绿素最大值层分布及其对悬浮体粒度的影响. 海洋与湖沼, 48(6): 1404-1414 |
甘雨, 2021. 西太平洋卡罗琳海脊海山粗糙度、多尺度特征及类比研究[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 28.
|
杨作升, 李云海, 2007. 太平洋悬浮体特征及近底雾状层(雾浊层)探讨. 海洋学报, 29(2): 74-81 |
杨作升, 郭志刚, 王兆祥, 等, 1992. 黄、东海水体中的有机包膜及其沉积作用. 海洋与湖沼, 23(2): 222-226 |
高微, 2018. 西太平洋暖池区北部悬浮体分布特征及其影响机制[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 65.
|
高微, 王珍岩, 张凯南, 等, 2016. 中尺度涡控制悬浮体的分布——以帕里西维拉海盆区为例. 海洋与湖沼, 47(4): 689-695 |
BAZZANO A, RIVARO P, SOGGIA F, et al, 2014. Anthropogenic and natural sources of particulate trace elements in the coastal marine environment of Kongsfjorden, Svalbard. Marine Chemistry, 163: 28-35 DOI:10.1016/j.marchem.2014.04.001 |
BISHOP J K B, COLLIER R W, KETTENS D R, et al, 1980. The chemistry, biology, and vertical flux of particulate matter from the upper 1500 m of the Panama Basin. Deep Sea Research Part A. Oceanographic Research Papers, 27(8): 615-640 DOI:10.1016/0198-0149(80)90077-1 |
BROTAS V, BREWIN R J W, SÁ C, et al, 2013. Deriving phytoplankton size classes from satellite data: Validation along a trophic gradient in the eastern Atlantic Ocean. Remote Sensing of Environment, 134: 66-77 DOI:10.1016/j.rse.2013.02.013 |
CHO B C, AZAM F, 1990. Biogeochemical significance of bacterial biomass in the ocean's euphotic zone. Marine Ecology Progress Series, 63(2/3): 253-259 |
DAI S, ZHAO Y F, LI X G, et al, 2020. The seamount effect on phytoplankton in the tropical western Pacific. Marine Environmental Research, 162: 105094 DOI:10.1016/j.marenvres.2020.105094 |
DJURHUUS A, READ J F, ROGERS A D, 2017. The spatial distribution of particulate organic carbon and microorganisms on seamounts of the South West Indian Ridge. Deep -Sea Research II: Topical Studies in Oceanography, 136: 73-84 DOI:10.1016/j.dsr2.2015.11.015 |
DOWER J, FREELAND H, JUNIPER K, 1992. A strong biological response to oceanic flow past Cobb Seamount. Deep Sea Research Part A. Oceanographic Research Papers, 39(7/8): 1139-1145 |
GAO W, WANG Z Y, LI X G, et al, 2021. The increased storage of suspended particulate matter in the upper water of the tropical Western Pacific during the 2015/2016 super El Niñ o event. Journal of Oceanology and Limnology, 39(5): 1675-1689 DOI:10.1007/s00343-021-0362-0 |
GENIN A, BOEHLERT G W, 1985. Dynamics of temperature and chlorophyll structures above a seamount: an oceanic experiment. Journal of Marine Research, 43(4): 907-924 DOI:10.1357/002224085788453868 |
HOGG N G, 1973. On the stratified Taylor column. Journal of Fluid Mechanics, 58(3): 517-537 DOI:10.1017/S0022112073002302 |
HU D X, WU L X, CAI W J, et al, 2015. Pacific western boundary currents and their roles in climate. Nature, 522(7556): 299-308 DOI:10.1038/nature14504 |
KANEKO I, TAKATSUKI Y, KAMIYA H, 2001. Circulation of intermediate and deep waters in the Philippine Sea. Journal of Oceanography, 57(4): 397-420 DOI:10.1023/A:1021565031846 |
KASHINO Y, UEKI I, SASAKI H, 2015. Ocean variability east of Mindanao: mooring observations at 7°N, revisited. Journal of Geophysical Research: Oceans, 120(4): 2540-2554 DOI:10.1002/2015JC010703 |
LAVELLE J W, MOHN C, 2010. Motion, commotion, and biophysical connections at deep ocean seamounts. Oceanography, 23(1): 90-103 DOI:10.5670/oceanog.2010.64 |
LE BORGNE R, BARBER R T, DELCROIX T, et al, 2002. Pacific warm pool and divergence: temporal and zonal variations on the equator and their effects on the biological pump. Deep Sea Research Part II: Topical Studies in Oceanography, 49(13/14): 2471-2512 |
LI W J, WANG Z Y, HUANG H J, 2020. Indication of size distribution of suspended particulate matter for sediment transport in the South Yellow Sea. Estuarine, Coastal and Shelf Science, 235: 106619 DOI:10.1016/j.ecss.2020.106619 |
LI W J, WANG Z Y, LEE G H, et al, 2021. Spatial variation of and the factors influencing the floc size distribution in the North Yellow Sea during the winter season. Marine Geology, 442: 106660 DOI:10.1016/j.margeo.2021.106660 |
LUKAS R, YAMAGATA T, MCCREARY J P, 1996. Pacific low-latitude western boundary currents and the Indonesian throughflow. Journal of Geophysical Research: Oceans, 101(C5): 12209-12216 DOI:10.1029/96JC01204 |
MA J, SONG J M, LI X G, et al, 2019. Environmental characteristics in three seamount areas of the Tropical Western Pacific Ocean: Focusing on nutrients. Marine Pollution Bulletin, 143: 163-174 DOI:10.1016/j.marpolbul.2019.04.045 |
MA J, SONG J M, LI X G, et al, 2021a. Multidisciplinary indicators for confirming the existence and ecological effects of a Taylor column in the Tropical Western Pacific Ocean. Ecological Indicators, 127: 107777 DOI:10.1016/j.ecolind.2021.107777 |
MA J, SONG J M, LI X G, et al, 2021b. Seawater stratification vs. plankton for oligotrophic mechanism: A case study of M4 seamount area in the Western Pacific Ocean. Marine Environmental Research, 169: 105400 DOI:10.1016/j.marenvres.2021.105400 |
MARTIN J, TREMBLAY J É, GAGNON J, et al, 2010. Prevalence, structure and properties of subsurface chlorophyll maxima in Canadian arctic waters. Marine Ecology Progress Series, 412: 69-84 DOI:10.3354/meps08666 |
MESSIÉ M, RADENAC M H, 2006. Seasonal variability of the surface chlorophyll in the western tropical Pacific from SeaWiFS data. Deep Sea Research Part I: Oceanographic Research Papers, 53(10): 1581-1600 DOI:10.1016/j.dsr.2006.06.007 |
MILLIMAN J D, FARNSWORTH K L, 2013. River Discharge to the Coastal Ocean: A Global Synthesis. Cambridge: Cambridge University Press
|
MURRAY J W, BARBER R T, ROMAN M R, et al, 1994. Physical and biological controls on carbon cycling in the equatorial Pacific. Science, 266(5182): 58-65 DOI:10.1126/science.266.5182.58 |
QU T D, LINDSTROM E J, 2004. Northward intrusion of Antarctic intermediate water in the western pacific. Journal of Physical Oceanography, 34(9): 2104-2118 DOI:10.1175/1520-0485(2004)034<2104:NIOAIW>2.0.CO;2 |
READ J, POLLARD R, 2017. An introduction to the physical oceanography of six seamounts in the southwest Indian Ocean. Deep-Sea Research II: Topical Studies in Oceanography, 136: 44-58 DOI:10.1016/j.dsr2.2015.06.022 |
ROGERS A D, 1994. The biology of seamounts. Advances in Marine Biology, 30: 305-350 |
SHI X Y, WANG Z Y, HUANG H J, 2021. Physical oceanography of the Caroline M4 seamount in the tropical Western Pacific Ocean in summer 2017. Journal of Oceanology and Limnology, 39(5): 1634-1650 DOI:10.1007/s00343-021-0359-8 |
TSUCHIYA M, 1991. Flow path of the Antarctic intermediate water in the western equatorial South Pacific Ocean. Deep Sea Research Part A. Oceanographic Research Papers, 38(Suppl 1): S273-S279 |
TURNER J T, 2015. Zooplankton fecal pellets, marine snow, phytodetritus and the ocean's biological pump. Progress in Oceanography, 130: 205-248 DOI:10.1016/j.pocean.2014.08.005 |
TURNER A, MILLWARD G E, 2002. Suspended particles: their role in estuarine biogeochemical cycles. Estuarine, Coastal and Shelf Science, 55(6): 857-883 DOI:10.1006/ecss.2002.1033 |
United States Board on Geographic Names, 1981. Gazetteer of Undersea Features: Names Approved by the United States Board on Geographic Names. 3rd ed. Washington: Defense Mapping Agency
|
WANG F, ZANG N, LI Y X, et al, 2015. On the subsurface countercurrents in the Philippine Sea. Journal of Geophysical Research: Oceans, 120(1): 131-144 DOI:10.1002/2013JC009690 |
WANG Z Y, SHI X Y, HUANG H J, 2022. Observation of physical oceanography at the Y3 seamount (Yap Arc) in winter 2014. Journal of Oceanology and Limnology, 40(4): 1314-1332 |
WHITE M, BASHMACHNIKOV I, ARISTEGUI J, et al, 2007. Physical processes and seamount productivity [M] //PITCHER T J, MORATO T, HART P J B, et al. Seamounts: Ecology, Fisheries & Conservation. The United Kingdom: Blackwell Publishing: 65-84.
|
WRIGHT D J, LODER J W, 1985. A Depth-dependent Study of the Topographic Rectification of Tidal Currents. Geophysical Astrophysical Fluid Dynamics, 31(3-4): 169-220 |
YESSON C, LETESSIER T B, NIMMO-SMITH A, et al, 2021. Improved bathymetry leads to > 4000 new seamount predictions in the global ocean-but beware of phantom seamounts!. UCL Open Environment, 4(3) DOI:10.14324/111.444/ucloe.000030 |
ZHANG L L, HU D X, HU S J, et al, 2014. Mindanao Current/Undercurrent measured by a subsurface mooring. Journal of Geophysical Research: Oceans, 119(6): 3617-3628 |
ZHANG L L, WANG F J, WANG Q Y, et al, 2017. Structure and variability of the north equatorial current/undercurrent from mooring measurements at 130°E in the western pacific. Scientific Reports, 7: 46310 |
ZHAO Y C, ZHAO Y, ZHENG S, et al, 2020. Virioplankton distribution in the tropical western Pacific Ocean in the vicinity of a seamount. MicrobiologyOpen, 9(6): 1207-1224 |