 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 严峻, 徐志进, 李铁军, 王慧君, 张冰妍, 钱红. 2023.
- YAN Jun, XU Zhi-Jin, LI Tie-Jun, WANG Hui-Jun, ZHANG Bing-Yan, QIAN Hong. 2023.
- 浙江舟山近海仔稚鱼时空动态变化及与环境因子的关系
- SPATIOTEMPORAL DYNAMICS OF LARVAE AND JUVENILES IN ZHOUSHAN COASTAL WATERS AND THEIR RELATIONSHIP WITH ENVIRONMENTAL FACTORS
- 海洋与湖沼, 54(3): 799-810
- Oceanologia et Limnologia Sinica, 54(3): 799-810.
- http://dx.doi.org/10.11693/hyhz20221000282
文章历史
-
收稿日期:2022-10-31
收修改稿日期:2023-02-19




2. 浙江省舟山市水产研究所 浙江舟山 316111;
3. 浙江海洋大学 浙江舟山 316022;
4. 浙江省海洋生态环境监测中心 浙江舟山 316021
2. Zhejiang Zhoushan Institute of Fisheries, Zhoushan 316111, China;
3. Zhejiang Ocean University, Zhoushan 316022, China;
4. Zhejiang Marine Ecology and Environment Monitoring Center, Zhoushan 316021, China
仔稚鱼既是微型浮游动物的捕食者, 又是很多海洋生物的饵料(于雯雯等, 2017), 仔稚鱼代表了海洋食物链中的一个重要营养环节(卞晓东, 2010)。鱼卵和仔稚鱼是鱼类的早期发育阶段, 对渔业资源的补充具有重要生态学意义(李建生等, 2007; 万瑞景等, 2014)。环境理化因子如水温、盐度等对于鱼卵和仔稚鱼的分布和存活具有直接和间接的影响。有研究指出, 环境理化因子的变动对河口区产卵的鱼类的后代数量影响最为剧烈和明显(李建生等, 2007)。不同类型的仔稚鱼必须广泛适应这一环境特点才能生存发育, 首先是对温度和盐度的适应性最为明显。
舟山群岛地处著名的舟山渔场, 是东海区恋礁性和洄游性鱼类的重要栖息场所, 在海洋动力环境上主要受长江冲淡水、浙北沿岸流和台湾暖流的影响。而近岸海域受长江冲淡水和浙北沿岸流影响较大, 海域水文环境独特, 饵料生物丰富、生物资源种类繁多, 是多种海洋经济鱼类以及各种水生生物产卵和索饵场所, 也是鱼类入海或溯河洄游的通道, 主产大黄鱼(Larimichthys crocea)、小黄鱼(Larimichthys polyactis)、带鱼(Trichiurus japonicus)等, 历史上曾是我国重要的海洋渔业资源分布区(樊紫薇等, 2020)。自20世纪60年代后期开始, 由于渔业资源的过度捕捞、海洋环境污染及气候变化等诸多因素, 使得舟山群岛渔业资源衰退日趋严重(李增光, 2013), 产卵场和栖息地被破坏, 影响了鱼类产卵环境, 进而影响仔稚鱼的集群结构(李圣法, 2005)。
近年来, 国内外关于仔稚鱼的研究较多, 主要关注环境因子对仔稚鱼的影响以及对成鱼的补充(Chen et al, 2014; 万瑞景等, 2014; 胡艳, 2015; Guan et al, 2017; Lin et al, 2018; 李建生等, 2018; 蒋日进等, 2019; 樊紫薇等, 2020), 包括种类组成、数量时空分布、多样性特征及其与环境因子(温度、盐度)的关系等。舟山群岛海域是我国传统的“三场一通道”(产卵场、索饵场、越冬场和洄游通道), 但目前的调查发现这些栖息地发生了变化, 高值的经济鱼种(小黄鱼、大黄鱼等)仔稚鱼出现概率较低, 其他低值鱼种(龙头鱼、虾虎鱼等)仔稚鱼出现率明显增多。仔稚鱼出现率的变化反映了环境的变化或人类活动的影响, 目前这方面的研究未见报道。
本文利用GLM模型建立了不同生态类群出现概率与环境因子之间的定量模型, 全面分析了仔稚鱼出现率、群落结构季节动态变化的主要变化机制, 旨在探明舟山近海海域仔稚鱼丰度的分布、出现概率受到种类本身及水文和环境因素(温度、盐度)的综合关系, 为进一步研究舟山群岛海域渔业资源的补充机制及生态修复和制定可持续发展措施提供科学依据。
1 材料与方法 1.1 调查站位及样品采集于2019年4月(春季)、7月(夏季)、11月(秋季)、3月(冬季)在舟山近海海域开展了4个航次的仔稚鱼调查。调查区域为29°~32°N, 121°~125°E, 分别设置站位31个、24个、46个、32个(图 1), 近岸平均水深约8~10 m, 近海约15~60 m。
 |
| 图 1 舟山近海2019年春(a)、夏(b)、秋(c)、冬(d)四个季节调查取样站位分布图 Fig. 1 Distribution of sampling stations in spring (a), summer (b), autumn (c), and winter (d) of 2019 in Zhoushan offshore |
仔稚鱼的采集采用Ⅰ型浮游生物网(口径500 mm, 网长1 450 mm, 0.505 mm)逐站进行表层水平拖网采样, 每站拖10 min, 拖速为2~3 n mile/h, 对温度、盐度等相关环境因子数据采用温盐深仪进行同步采集。采集样品用5%福尔马林溶液固定, 带回实验室, 再根据仔稚鱼的形态特征进行种类鉴定, 并计数。仔稚鱼样品的采集、保存、运输和分析均按照《海洋调查规范—海洋生物调查》(GB/T 12763.6-2007)中的规定执行。
1.2 数据处理与分析方法 1.2.1 仔稚鱼类群划分参考相关文献(卞晓东, 2010; 万瑞景等, 2014; 肖欢欢等, 2017), 对仔稚鱼种类的生态类型和适温类型进行了划分。适温类型包括暖温种(也称广温广盐鱼类, 生存水温范围0~35 ℃)、暖水种(耐高温鱼类, 生存水温范围20~40 ℃)和冷温种(耐低温鱼类, 生存水温范围0~20 ℃), 生态类型划分为河口型(包括河口类, 生活在河口附近, 长期生活在河口盐度在5~15的中盐性海域)、沿岸型(春、夏季由较深的越冬海区游至河口附近的浅海或者近河口的半咸水区生殖, 生殖后的鱼群及大量的幼鱼在此索饵育肥)和近海型(主要是季节性洄游种)。在本研究中, 将河口型仔稚鱼与沿岸型仔稚鱼统归为近岸河口型进行模型构建。为降低仔稚鱼数据的极化程度, 减少稀有种的影响, 对仔稚鱼尾数采用对数转化:
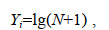 (1)
(1)式中, N为原始尾数, Yi为转换后数值。
1.2.2 仔稚鱼丰度仔稚鱼丰度采用公式(2)计算:
 (2)
(2)式中, N为单位体积海水中的仔稚鱼个体数(ind./m3); V为采集仔稚鱼样品过滤海水体积(m3); n为样品鉴定计数获得的仔稚鱼个体数; r为采样网口半径, l为拖网深度或者拖网距离。
1.2.3 生态优势度仔稚鱼生态优势度采用公式(3)计算:
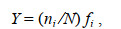 (3)
(3)式中, N为各采样站位所有种类的尾数; ni为第i种的尾数; fi为该种在各站位出现的频率。根据以往学者的观点, 本文将Y > 0.02的种类定为优势种。环境因子空间分布、仔稚鱼类群丰度分布图等均使用Sufer11.0绘制。
1.2.4 GLM模型的应用GLM是一般线性模型扩展, 由Nelder和Wed-derburn在1972年提出。一般线性模型是描述一个或多个自变量之间的线性依存关系。它要求因变量连续且服从正态分布, 通过自变量的线性预测值得到因变量估计值。但在实际应用中, 有很多数据并不能完全满足一般线性模型的应用条件(尹洁等, 2019)。因此, 为了进一步扩展其应用, 通过对应变量Y的期望值进行非线性函数转化, 发展出了GLM模型, 一般形式为:
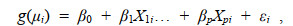 (4)
(4)式中, 因变量相互独立, 服从指数分布族(如正态分布、二项分布、泊松分布等), 因变量的期望值为E(yi)=μi; β0为截距; β1X1i… + βpXpi为线性部分, β为偏回归系数, X为自变量; g(.)为联接函数, 它将因变量的期望值和线性部分联接起来, 其作用是对因变量进行变换, 使其符合线性模型的条件(曾韦霖等, 2020)。
1.3 仔稚鱼出现概率模型的构建由于近岸河口型仔稚鱼、近海型仔稚鱼在春秋季有不同的最适温度, 本研究假设温度和盐度对仔稚鱼的出现具有显著影响, 并利用这些预测变量来定义其栖息地的模型。具体来说, 通过构建栖息地模型, 来预测仔稚鱼的出现概率。出现概率模型使用了2019年春、秋季从舟山群岛附近的77个采样站位收集的数据(图 1)。
采用SPSS(2019版)进行logistic回归分析。用logistic回归分析了近岸河口型仔稚鱼与近海型仔稚鱼的存在与温度和盐度之间的关系, 以估计其出现概率。Logistic回归在将因变量(仔稚鱼的存在、仔稚鱼的缺失)转换为Logit变量后应用最大似然估计, 其中Logit是仔稚鱼出现几率的自然对数。Logistic回归是一个合适的模型, 因为它将一个二元因变量(存在/不存在)与连续或分类的自变量进行回归, 放宽了自变量正态性的假设(Decker et al, 2007)。
1.4 自变量的选择使用相关性分析(correlation analysis)和逐步回归(stepwise regression)选取自变量和交互项, 使用AIC (akaike information criterion)准则检验模型的适合程度, 选择AIC值小的模型为最优模型。AIC值由下式计算:
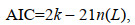 (5)
(5)式中, k是参数的个数, n是样本数量, L是似然函数(李增光, 2013)。
1.5 模型验证为了评估经验推导的栖息地模型的预测性能, 本研究针对独立的仔稚鱼和环境数据测试了近岸鱼类的出现概率模型。模型预测是通过比较这些模型衍生的事件与测试数据集中观察到的仔稚鱼数量来评估的(Decker et al, 2007)。
2 结果 2.1 环境因子杭州湾和舟山近海在不同季节的SST (sea surface temperature, 海表温度)、SSS (sea surface salinity, 海表盐度)均呈现出近岸低、近海高的趋势(图 2)。2019年平均温度7月(25.59 ℃) > 11月(20.76 ℃) > 4月(15.44 ℃) > 3月(11.63 ℃), 平均盐度3月(30.33) > 11月(26.40) > 7月(25.16) > 4月(24.49)。
 |
| 图 2 2019年春、夏、秋、冬季舟山近海海域表层温度(SST)、盐度(SSS)分布与季节变化 Fig. 2 Distribution and monthly variation of surface temperature (SST, a~d) and salinity (SSS, e~h) in Zhoushan coastal waters in spring, summer, autumn, and winter of 2019 注: a. 春季SST, b. 夏季SST, c. 秋季SST, d. 冬季SST, e. 春季SSS, f. 夏季SSS, g. 秋季SSS, h. 冬季SSS |
2019年舟山群岛及邻近海域4个季度共采集仔稚鱼228尾(表 1)。经鉴定分析共有仔稚鱼28种, 隶属于9目18科28属, 另有未定种4种; 鲈形目种类最多, 有9科17种(60.71%); 其次是鲱形目, 有1科3种(10.71%)。仔稚鱼种类数: 春季(19种) > 夏季(16种) > 秋季(10种) > 冬季(5种), 四季共有科仅虾虎鱼科、石首鱼科、灯笼鱼科3种, 显示仔鱼种类有明显的季节性变化。
| 种类 | 适温性、生态类型 | 春季 | 夏季 | 秋季 | 冬季 |
| 鲈形目Perciformes | |||||
| 虾虎鱼科Gobiidae | + | + | |||
| 斑尾刺虾虎鱼Acanthogobius ommaturus | E & WT | + | + | ||
| 阿部鲻虾虎鱼Mugilogobius abei | E & WT | + | |||
| 纹缟虾虎鱼Tridentiger trigonocephalus | E & WT | + | |||
| 六丝钝尾虾虎鱼Amblychaeturichthys hexanema | E & WT | + | + | ||
| 舌虾虎鱼Glossogobius giuris | E & WW | + | |||
| 拟矛尾虾虎鱼Parachaeturichthys polynema | C & WT | + | |||
| 拉氏狼牙虾虎鱼Odontamblyopus lacepedii | E & WT | + | + | ||
| 石首鱼科Sciaenidae | + | ||||
| 小黄鱼Larimichthys polyactis | C & WT | + | + | ||
| 大黄鱼Larimichthys crocea | C & WT | + | |||
| 黄姑鱼Nibea albiflora | C & WW | + | |||
| 白姑鱼Argyrosomus argentatus | C & WW | + | |||
| 带鱼科Trichiurid | |||||
| 狭颅带鱼Tentoriceps cristatus | C & WW | + | |||
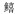 科Sillaginidae 科Sillaginidae |
|||||
| 少鳞鱚Sillago japonica | N & WW | + | |||
| 鲭科Scombridae | |||||
| 日本鲭Scomber japonicus | N & WW | + | |||
| 鲳科Stromateidae | |||||
| 银鲳Pampus argenteus | N & WT | + | |||
| 鮨科Serranidae | |||||
| 花鲈Lateolabrax japonicus | E & WT | + | |||
 科Callionymidae 科Callionymidae |
|||||
| 香斜棘 Repomucenus olidus |
C & WW | + | |||
| 鲷科Sparidae | + | ||||
| 鲽形目Pleuronectiformes | |||||
| 舌鳎科Cynoglossidae | + | ||||
| 舌鳎属Cynoglossus | + | ||||
| 鲽科Pleuronectidae | + | ||||
| 鲉形目Scorpaeniformes | |||||
| 鲉科Scorpaenidae | + | + | |||
| 褐菖鲉Sebastiscus marmoratus | N & WW | + | |||
| 红娘鱼属Lepidotrigla | + | ||||
| 鲱形目Clupeiformes | |||||
| 鳀科Engraulidae | |||||
| 中华小公鱼Stolephorus chinensis | N & WW | + | + | + | |
| 凤鲚Coilia mystus | E & WT | + | |||
| 日本鳀Engraulis japonicus | N & WT | ||||
| 仙女鱼目Aulopiformes | |||||
| 合齿鱼科Synodontidae | |||||
| 龙头鱼Harpadon nehereus | C & WT | + | + | ||
| 鲤形目Cypriniformes | |||||
| 鲤科Cyprinidae | |||||
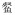 Hemiculter leucisculus Hemiculter leucisculus |
E & WT | + | |||
| 灯笼鱼目Myctophiformes | |||||
| 灯笼鱼科Myctophidae | + | + | |||
| 七星底灯鱼Benthosema pterotum | N & WW | + | + | ||
| 鲻形目Mugiliformes | |||||
| 鲻科Mugilidae | + | ||||
| 鲻鱼Mugil cephalus | C & WT | + | |||
| 鲑形目Salmoniformes | |||||
| 银鱼科Salangidae | + | ||||
| 未定种 | |||||
| 未定种仔鱼1 | + | + | |||
| 未定种仔鱼2 | + | + | |||
| 注: E & WT: 河口暖温种; E & WW: 河口暖水种; N & WW: 近海暖水种; N & WT: 近海暖温种; C & WW: 沿岸暖水种; C & WT: 沿岸暖温种 | |||||
按种类适温属性划分, 调查海区仅出现暖温性和暖水性两种, 其中暖温性15种(占已知适温类型种类的60%), 暖水性10种(占40%); 按生态类型划分主要包括河口型(9种, 36%)、沿岸型(9种, 36%)、近海型(7种, 28%)。
2.2.2 优势种组成舟山群岛海域仔稚鱼优势种的组成存在一定的季节差异性。春季, 出现仔稚鱼优势种1种, 为暖温性鱼类斑尾刺虾虎鱼(44尾, 45.10%); 夏季, 仔稚鱼优势种仅1种, 为暖温性鱼类拉式狼牙虾虎鱼(28尾, 53.85%); 秋季, 仔稚鱼优势种有2种, 分别为侧带小公鱼(23尾, 41.82%)和银鱼科(14尾, 25.45%); 冬季, 仔稚鱼优势种有2种, 分别为鲷科(7尾, 35%)和灯笼鱼科(7尾, 35%)。全年仔稚鱼优势种有四种, 分别是斑尾刺虾虎鱼、侧带小公鱼属、拉氏狼牙虾虎鱼、六丝钝尾虾虎鱼(表 2)。
| 月份 | 优势物种 | 优势度 | 丰度比例/% |
| 春季 | 斑尾刺虾虎鱼Acanthogobius ommaturus | 0.131 | 45.10 |
| 夏季 | 拉式狼牙虾虎鱼Odontamblyopus lacepedii | 0.377 | 53.85 |
| 秋季 | 侧带小公鱼Stolephorus chinensis、银鱼科Salangidae | 0.134 | 41.82 |
| 0.051 | 25.45 | ||
| 冬季 | 灯笼鱼科Myctophidae、鲷科Sparidae | 0.073 | 35.00 |
| 0.058 | 35.00 | ||
| 全年 | 斑尾刺虾虎鱼Acanthogobius ommaturus | 0.107 | 21.49 |
| 侧带小公鱼Stolephorus chinensis | 0.089 | 11.84 | |
| 拉式狼牙虾虎鱼Odontamblyopus lacepedii | 0.064 | 12.72 | |
| 六丝钝尾鰕虎鱼Amblychaeturichthys hexanema | 0.024 | 4.82 |
从总丰度来看, 4个航次的调查中, 各航次调查区域仔稚鱼密度分布近岸高, 近海低(图 3)。自春季至冬季, 仔稚鱼密度呈先增后减的趋势, 其中夏季航次各站点仔稚鱼平均密度最大0.270 ind./m3, 其次是春季0.007 ind./m3, 最后是秋季和冬季都为0.003 ind./m3; 四个季节近岸河口型仔稚鱼平均密度为0.023 ind./m3, 近海型仔稚鱼平均密度为0.011 ind./m3。
 |
| 图 3 2019年春、夏、秋、冬季仔稚鱼物种分布图 Fig. 3 Species distribution of larvae and juveniles in spring, summer, autumn, and winter of 2019 注: 仔稚鱼总丰度分布: a. 春季, b. 夏季, c. 秋季, d. 冬季; 近岸河口型仔稚鱼丰度分布: e. 春季, f. 夏季, g. 秋季, h. 冬季; 近海型仔稚鱼丰度分布: i. 春季, j. 夏季, k. 秋季, l. 冬季 |
从分布区域来看, 春季仔稚鱼集中在杭州湾及近岸海域, 夏季仔稚鱼分布海区整体向北移动, 在长江口冲淡区密集分布, 秋季仔稚鱼分布向长江口南部及舟山近海海域移动, 冬季仔稚鱼无明显聚集区, 仅在近岸与近海交界带形成一个较小的密集带。
2.3 仔稚鱼的出现概率模型构建本研究发现仔稚鱼的丰度与温度和盐度有着密切的关系, 以下方程分别预测了两种生态类型仔稚鱼出现的概率与温度与盐度的关系:
近岸河口型仔稚鱼:
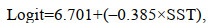 (6)
(6)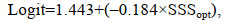 (7)
(7)近海型仔稚鱼:
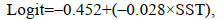 (8)
(8)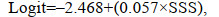 (9)
(9)式中, Logit是仔稚鱼出现概率的自然对数, SSSopt是最佳盐度(optimum salinity)。
S型概率曲线通过转换每个线性回归函数来拟合温度和盐度数据:
 (10)
(10)式中, Pocc是仔稚鱼出现的概率(图 4)。可以发现与最佳盐度之间的差异变小的趋势与近岸河口型仔稚鱼出现相对高的概率有关, 并且, 随着最佳盐度与实际盐度之间的差异的增加, 这种出现概率下降。
 |
| 图 4 近岸河口型仔稚鱼出现的概率与海面温度之间的关系(a), 与最佳盐度之间差异的关系(b); 近海型仔稚鱼出现的概率与海面温度之间的关系(c), 与盐度之间的关系(d) Fig. 4 Relationships of the probability of larvae-and-juveniles occurrence in inshore estuary to the sea surface temperature (a), and to the difference in optimal salinity (b); relationship of the probability of the occurrence in offshore to the sea surface temperature (c) and to the salinity (d) 注: 温度和盐度都是仔稚鱼存在的良好预测因子, 趋势线由逻辑回归分析确定 |
为了建立一个同时考虑温度和盐度的仔稚鱼出现概率模型, 在本研究中采用多元logistic回归模型分析了这些变量之间的关系。Hosmer-lemeshow test将包括温度和盐度在内的完整模型与单因素的简化模型进行比较, 结果表明, 近岸鱼类出现的概率是温度和盐度的函数: Logit=8.138+(–0.373×SST)+(–0.169× SSSopt), 包括温度和盐度的多元logistic回归比包含温度(df=1, P < 0.001)或盐度(df=1, P < 0.001)的简化单变量方程更适合数据。近海型仔稚鱼出现的概率是盐度的函数: Logit=–2.468+(–0.057×SSS), 仅包括盐度(df=1, P=0.175 > 0.05)的简化单变量方程比包含温度和盐度的多元logistic回归更适合数据。两个函数的Hosmer-Leme拟合优度检验分别为13.256、5.265, 不显著(P > 0.05), 表明模型与数据拟合良好。
2.4 仔稚鱼出现概率与各因子的关系使用逐步回归, 以AIC值最小的为最优模型, 可以看出GLM模型各因子对仔稚鱼出现概率产生的影响(表 3), 从表 3可看出, 对于近岸河口型仔稚鱼, 温度和盐度差值为最大, 其次是温度和盐度差值和温度与盐度差值的交互效应, 对于近海型仔稚鱼, 盐度为最大, 其次是温度和盐度。近岸河口型仔稚鱼选择了表层温度、最佳盐度作为最优模型的环境变量, 近海型仔稚鱼选择了表层盐度最优模型的环境变量。
| 仔稚鱼类型 | 模型 | P | AIC |
| 近岸河口型仔稚鱼 | Inshore: SST | P<0.05*** | 89.02 |
| Inshore: SSSopt | P<0.05*** | 91.037 | |
| Inshore: SST + SSSopt | P<0.05** | 80.344 | |
| Inshore: SST + SSSopt + SST×SSSopt | P>0.05 | 82.237 | |
| 近海型仔稚鱼 | Offshore: SST | P>0.05 | 94.135 |
| Offshore: SSS | P>0.05 | 92.242 | |
| Offshore: SST+SSS | P>0.05 | 93.307 | |
| Offshore: SST + SSS + SST×SSS | P>0.05 | 94.802 | |
| 注: ***表示P<0.001; **表示P<0.01; *表示P<0.05 | |||
作者拟通过实际采集的仔稚鱼数据与本研究的模型预测结果数据进行分析, 以评估本研究预测的可靠性。以2018年调查数据为实例, 实际采样所得的近岸河口型仔稚鱼、近海型仔稚鱼均基本出现在模型预测概率高值区(图 5)。从季节方面来看, 春季模型较秋季模型更符合预期; 从生态类群来看, 近岸河口型仔稚鱼模型预测效果较近海仔稚鱼更符合预期。可见本次研究的预测模型对预测仔稚鱼的出现概率能起到一定的作用。
 |
| 图 5 2018年春季近岸河口型仔稚鱼(a)、近海型仔稚鱼(b); 2018年秋季近岸河口型仔稚鱼(c)、近海型仔稚鱼(d)出现概率预测与实际丰度验证图 Fig. 5 Prediction of occurrence probability and verification of actual abundance of coastal estuarine larvae and juveniles (a), offshore larvae and juveniles (b) in spring 2018; coastal estuarine larvae and juveniles (c), offshore larvae and juveniles (d) in autumn 2018 注: “+”表示出现概率预测; 黑点表示实际丰度 |
本研究模型表明, 近岸河口型仔稚鱼和近海型仔稚鱼的栖息地具有狭义的温度和盐度组合, 这种对特定热和盐分条件的“偏好”决定了舟山近海海域仔稚鱼分布的时空格局。据杨东莱等发现, 长江口的鱼类浮游生物中, 半咸水型(河口型)的多数种主要于春季水温为12.0~22.0 ℃、盐度为0.1~12.0的长江下游水道及其入海口附近海区进行繁殖, 沿岸型多分布在盐度为5.0~26.0的混浊水域, 即122°30’E以西、25 m等深线以内的浅水区, 近海型主要在盐度24.0~ 33.0、水质澄清的海区, 即122°30’E以东、30 m等深线以外的海区(刘淑德, 2009)。本文将半咸水种(河口型)和沿岸种合并为近岸河口型进行讨论, 可以发现, 近岸河口型仔稚鱼存在温度阈值12 ℃。近海型仔稚鱼存在盐度阈值33.0, 本研究结果与前人研究结果一致(丁月旻等, 2011)。
仔稚鱼的分布不仅受成体繁殖习性的影响, 而且也与环境因子分布密切相关。在本研究中, 两种生态类群仔稚鱼的分布特征都验证了这一观点。春季是大多数鱼类的产卵季节, 舟山近海由于具有高浑浊度、丰富的有机碎屑, 是传统的产卵场与育幼场。自春季(4月)开始, 长江冲淡水的强度和范围增大(要津, 2017), 舟山海域水温和盐度等物理环境因子差异明显, 形成了较为明显的温度锋面, 仔稚鱼多分布于温度锋面温度偏高一侧, 这一对高温、高浑浊度海域的喜好可能反映了仔稚鱼的生殖策略。同时, 本研究发现近海型仔稚鱼具有较强的环境适应性和灵活性。在春季, 舟山近海海域的表温逐步上升, 仔稚鱼的丰度也逐渐上升, 以日本鳀鱼为例, 当4月表层水温达到13 ℃ (底层水温达到15 ℃)时, 鳀鱼开始进入沿岸产卵场产卵, 当水温达20 ℃以上时, 进入产卵盛期。产卵场表层水温范围13~24 ℃, 表层盐度23.13~ 31.88 (周永东等, 2013), 此温盐范围与日本鳀鱼丰度高值区采集的温盐数据相一致。在秋季, 舟山近海海域的表温整体趋于均匀温暖, 因此不会对仔稚鱼的出现产生显著的影响, 相比之下, 秋季盐度在舟山近海呈现近岸海域低、近海海域高的趋势, 该趋势对仔稚鱼在调查区域中的位置有着显著影响。仔稚鱼在整个调查海域的近岸、近海均下降, 这可能是由于在该季节随着台湾暖流势力的减弱, 一些鱼类游向外海深水区(郭朋军等, 2020)。历史研究也表明, 仔稚鱼群落结构存在着高度的时空异质性, 海洋地理特征的时空模式(如长江冲淡水、浙北沿岸流、台湾暖流)影响仔稚鱼聚集模式和生长, 从而影响渔业种群动态(卞晓东, 2010; 肖欢欢等, 2017)。
3.2 舟山海域的育幼场形成的潜在因素分析舟山近海的春、夏、秋、冬四个季节的仔稚鱼资源密度调查结果显示, 靠近杭州湾和长江口河口区的仔稚鱼丰度最高, 出现水域主要集中在受江浙沿岸水控制的低温低盐区; 在远离海岸而受混合水控制的高温高盐区仅有少量的分布(李建生等, 2007)。本次调查的优势类群为虾虎鱼科, 其产卵场主要分布在舟山近海北部的杭州湾内和舟山近海南部的鱼山渔场近岸水域, 从仔稚鱼时空分布图来看(图 6), 舟山近海南部的鱼山渔场是最早集中产卵的水域, 随着时间的推移, 产卵场的中心区逐渐向北部水域的舟山渔场转移(周永东等, 2011), 仔稚鱼丰度高值区也逐步从南往北转移, 这与调查水域的水温随时间从南到北逐渐升高有关, 舟山近海北部的杭州湾内产卵的鱼类主要是河口型, 属相对独立的产卵区域。仔稚鱼的分布与温度和盐度相关性显著, 这一结论在前人的大量报道中已经得到证实, 本文研究结果与前人研究结果一致(管伟, 2016; Guan et al, 2017)。
 |
| 图 6 2019年春(a)、夏(b)、秋(c)、冬(d)季仔稚鱼分布图 Fig. 6 Distribution of larvae and juveniles in spring (a), summer (b), autumn (c), and winter (d) of 2019 |
根据本文模型拟合结果, 温度对舟山海域的仔稚鱼的影响最为显著, 这主要是受鱼类产卵温度阈值比较小的影响, 直接关系到仔稚鱼的成活。同时, 除了温度影响外, 盐度对鱼卵的影响也很大, 长江淡水入海的影响、咸淡水的交汇给仔鱼带来适宜的盐度, 提高了成活率。本文结果为进一步研究舟山近海海域仔稚鱼丰度的分布和出现概率提供了基础。
丁月旻, 线薇微, 2011. 秋季长江口鱼类浮游生物群落时空结构. 中国海洋大学学报, 41(10): 67-74 |
于雯雯, 张东菊, 邹欣庆, 等, 2017. 海州湾海域浮游动物种类组成与丰度的季节变化. 生态学杂志, 36(5): 1339-1349 |
万瑞景, 曾定勇, 卞晓东, 等, 2014. 东海生态系统中鱼卵、仔稚鱼种类组成、数量分布及其与环境因素的关系. 水产学报, 38(9): 1375-1398 |
卞晓东, 2010. 鱼卵、仔稚鱼形态生态学基础研究——兼报黄河口海域鱼类浮游生物调查[D]. 青岛: 中国海洋大学.
|
尹洁, 王晶, 张崇良, 等, 2019. 利用two-stage GAM研究海州湾及其邻近海域小黄鱼鱼卵的时空分布特征. 中国水产科学, 26(6): 1164-1174 |
刘淑德, 2009. 长江口及其邻近海域鱼类浮游生物群落结构特征研究[D]. 青岛: 中国科学院研究生院(海洋研究所).
|
李圣法, 2005. 东海大陆架鱼类群落生态学研究—空间格局及其多样性[D]. 上海: 华东师范大学.
|
李建生, 胡芬, 程家骅, 2007. 长江口水域春季鱼卵仔鱼分布及其与温度盐度的关系. 海洋科学, 31(4): 16-21 |
李建生, 凌建忠, 胡芬, 2018. 长江口近岸水域小黄鱼仔稚鱼时空分布和生长特征. 海洋渔业, 40(4): 404-412 DOI:10.3969/j.issn.1004-2490.2018.04.003 |
李增光, 2013. 基于GAM模型的南黄海帆张网主要渔获物分布及海州湾鱼卵、仔稚鱼集群特征的初步研究[D]. 青岛: 中国海洋大学.
|
肖欢欢, 张崇良, 徐宾铎, 等, 2017. 黄海中南部近岸海域春季鱼类浮游生物群落空间格局研究. 海洋学报, 39(8): 34-47 |
周永东, 金海卫, 张洪亮, 等, 2013. 浙江南部沿岸产卵场春、夏季鱼卵、仔稚鱼种类组成与分布. 海洋与湖沼, 44(3): 590-598 |
周永东, 金海卫, 蒋日进, 等, 2011. 浙江中北部沿岸春、夏季鱼卵和仔稚鱼种类组成与数量分布. 水产学报, 35(6): 880-889 |
胡艳, 2015. 长江口近岸水域幼鱼时空分布及代表种资源评估[D]. 上海: 上海海洋大学.
|
要津, 2017. 春季长江口外上升流变化及其对悬浮体分布的影响[D]. 青岛: 中国科学院大学(中国科学院海洋研究所).
|
郭朋军, 江新琴, 俞存根, 等, 2020. 舟山沿岸渔场春秋季鱼类群落结构特征分析. 渔业科学进展, 41(4): 1-11 |
蒋日进, 张琳琳, 徐开达, 等, 2019. 浙江中南部近岸海域游泳动物功能群特征与多样性. 生物多样性, 27(12): 1330-1338 |
曾韦霖, 刘涛, 肖建鹏, 等, 2020. 广义线性模型在时间序列研究中的应用及R语言实现. 华南预防医学, 46(3): 321-323 |
管伟, 2016. 台湾海峡中部近海鱼类资源分布及群落结构的季节变化与水团的关系[D]. 上海: 上海海洋大学.
|
樊紫薇, 蒋日进, 张洪亮, 等, 2020. 岱衢洋产卵场鱼卵、仔稚鱼群落结构及其与环境因子的关系. 水产学报, 44(4): 606-620 |
CHEN W Y, LEE M A, LAN K W, et al, 2014. Distributions and assemblages of larval fish in the East China Sea during the northeasterly and southwesterly monsoon seasons of 2008. Biogeosciences, 11(3): 547-561 |
DECKER M B, BROWN C W, HOOD R R, et al, 2007. Predicting the distribution of the scyphomedusa Chrysaora quinquecirrha in Chesapeake Bay. Marine Ecology Progress Series, 329: 99-113 |
GUAN L, DOWER J F, MCKINNELL S M, et al, 2017. Interannual variability in the abundance and composition of spring larval fish assemblages in the Strait of Georgia (British Columbia, Canada) from 2007 to 2010. Fisheries Oceanography, 26(6): 638-654 |
LIN N, CHEN Y G, JIN Y, et al, 2018. Distribution of the early life stages of small yellow croaker in the Yangtze River estuary and adjacent waters. Fisheries Science, 84(2): 357-363 |