 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 梅泊承, 张晓东, 赵淳朴, 徐继林, 王丹丽, 郭春阳. 2023.
- MEI Bo-Cheng, ZHANG Xiao-Dong, ZHAO Chun-Pu, XU Ji-Lin, WANG Dan-Li, GUO Chun-Yang. 2023.
- 串联养殖模式下的凡纳滨对虾(Litopenaeus vannamei)与缢蛏(Sinonovacula constricta)生长特性、消化免疫及水生态效应
- GROWTH PERFORMANCE AND AQUATIC ECOLOGICAL EFFECT IN TANDEM CULTURE OF SHRIMP LITOPENAEUS VANNAMEI AND RAZOR CLAM SINONOVACULA CONSTRICTA
- 海洋与湖沼, 54(3): 907-920
- Oceanologia et Limnologia Sinica, 54(3): 907-920.
- http://dx.doi.org/10.11693/hyhz20221000280
文章历史
-
收稿日期:2022-10-29
收修改稿日期:2023-01-03




凡纳滨对虾(Litopenaeus vannamei)和缢蛏(Sinonovacula constricta)分别是我国东南沿海重要的海水养殖虾类和贝类, 养殖规模大, 产量高(薛宝宝, 2019; Yuan et al, 2020)。同时凡纳滨对虾养殖也在全球养殖数量中稳步上升。然而, 养虾产业迎来了不少的挑战, 近年来虾蛏海水围塘养殖存在着诸如生产模式效率低、养殖塘生态系统不稳定、物质与能量利用率不高、病害频发、养殖经济效益下滑等诸多问题, 已不能适应现代渔业可持续发展的需求(Nadella et al, 2021)。研究者针对传统这些问题, 进行不少研究和探索, 近年逐步形成了以内循环养殖为特征的池塘生态养殖模式, 比如运用在斑点叉尾鮰养殖中, 生产率较传统养殖有较好的提高(Brown et al, 2011)。这种模式的主要特点是将养殖池分为饲养生产区和水质净化区(Turker et al, 2003), 水产养殖系统分区是基于将浮游植物现存量生产力管理以增加鱼产量, 利用浮游植物的高生产力去除养殖对象释放的无机营养物(Smith et al, 1988; Drapcho et al, 2000)。本文所研究的凡纳滨对虾与缢蛏串联养殖模式也属于这种模式之一, 是将凡纳滨对虾与缢蛏分塘养殖的一种新模式(刘招坤, 2014; 张清科等, 2019), 依据凡纳滨对虾和缢蛏的不同生态位、习性和食性互补的特点, 有效降低水体中有害营养盐的积累, 维持水生态环境相对稳定, 实现虾蛏健康高效养殖。
实现健康高效养殖, 离不开微藻的调控。微藻是在海洋和淡水中发现的微小单细胞生物, 单独或成群生活在表面、沉积物中(曹豪豪等, 2021)。它们在食物链中起着重要的作用, 因其可以在不同的生命阶段被鱼类和贝类食用(王志滨等, 2014)。微藻除了吸收无机氮和磷等污染物之外, 还增加了养殖环境水体中有机碳含量, 这很好地改善了养殖水环境(Gupta et al, 2019)。另外, 微藻也含有抗氧化剂, 包括多不饱和脂肪酸、光合色素和类胡萝卜素酚类化合物等(Shalaby, 2011; Chen et al, 2021)。这些可能有助于对虾在紧张的水产养殖条件下减少活性化合物。随着食物链传递, 活性氧化物会在养殖动物体内逐渐积累, 可以减少氧化应激效应, 提高免疫性能, 但最终使得虾产量升高。
提高水产养殖的产量与合适的养殖密度关系密切。养殖密度在水产养殖中是一个重要参数, 对养殖对象生长性能有着明显影响。关于养殖密度研究方面, 多以鱼、虾、蟹为研究对象。较适的养殖密度不仅提高养殖成功率和减少病害发生, 而且也增加了养殖效益。但高密度养殖是水产养殖中很常见的一种慢性胁迫因子, 过高的养殖密度则会降低养殖对象生长率及消化酶酶活性(Huchette et al, 2003; Sahin et al, 2014)。陈志(2017)研究密度对双线紫贻幼贝生长存活影响, 发现高密度对贝生长率及存活率显著降低。近年, 骆云慧等(2021)、鲍格格等(2022)研究了虾蛏串养模式下的能量流动模型、浮游植物与水质之间的关系和生态修复效果, 但关于虾蛏串联养殖中两者不同的数量配比对虾蛏消化酶及非特异性免疫酶的影响方面未见相关研究报道。
养殖对象是否健康, 养殖环境是否友好, 这些不仅可以通过检测养殖对象的消化酶、非特异性免疫指标和养殖水环境的水质指标来综合判断, 也可以通过检测养殖对象及养殖环境微生物群落来评价。越来越多研究表明, 微生物在水产养殖生态系统中扮演着关键角色, 水体环境、底泥微生物有着氮磷循环、净化水质、污染物降解等功能(Newton et al, 2011; 任丽娟等, 2013); 而肠道微生物在宿主对摄入营养物质的消化吸收、生长发育、免疫等方面起着重要作用(金若晨, 2020), 例如, 作为抵抗病原体入侵的屏障并为宿主营养获取提供补充代谢途径(Crovesy et al, 2020; Zheng et al, 2020)。由于微生物群落易受到所处环境变化而变化, 如养殖对象数量的变化、营养物质输入、养殖对象排泄物和浮游生物残骸、季节水温变化及动保产品的使用等。这些因素都很有可能导致养殖水体、底泥及养殖对象中微生物群落组成及丰度改变, 从而影响到养殖对象的健康生长(Dai et al, 2022)。
本实验采用Illumina MiSeq平台对虾蛏不同数量配比串联养殖模式下的水体、底泥及缢蛏内脏团微生物的16S rRNA V3~V4区进行高通量测序, 比较并分析微生物群落组成及其与水环境因子的相互作用。并通过水环境因子变化、水体和底泥微生物群落结构等养殖外部因素, 虾蛏相关消化免疫酶指标及缢蛏内脏团微生物群落结构等内部因素综合分析, 确定虾蛏串养模式最适虾蛏养殖数量配比, 旨在为今后虾蛏串联养殖模式的高效健康发展提供科学依据。
1 材料与方法 1.1 实验材料健康缢蛏来自于宁波鄞州椿霖水产养殖场, 凡纳滨对虾来自于鄞州浩海水产养殖场。实验前暂养于天然海水(盐度20, 28~29 ℃)中。实验用蛏规格为(10.01±0.15) g, 凡纳滨对虾规格为(4.01±0.06) g。
1.2 方法 1.2.1 实验设计及养殖管理实验设置LP组、MP组、HP组3个虾蛏不同数量配比实验组, 每个实验组设3个平行组, 各实验组在对虾数量不变的情况下, 设定虾与蛏数量配比分别为1︰3、1︰5、1︰7, 具体投放虾蛏数量见表 1。实验时间为2021年9月30日~10月28日, 实验周期为28 d。
| 组别 | 虾密度/(ind./m2) | 蛏密度/(ind./m2) | 虾蛏比 | 对虾数量/尾 | 投放缢蛏数量/只 |
| LP组 | 60 | 180 | 1︰3 | 12 | 9 |
| MP组 | 60 | 300 | 1︰5 | 12 | 15 |
| HP组 | 60 | 420 | 1︰7 | 12 | 21 |
该实验在宁波大学中试基地的室内模拟虾蛏串联养殖系统中进行, 根据本实验室前期研究结果, 设定虾蛏养殖面积比为4︰1, 由一个面积为0.20 m2对虾养殖槽和一个面积为0.05 m2的缢蛏养殖槽组成一个独立的养殖单元。虾槽水深0.25 m, 蛏槽水深0.15 m, 缢蛏养殖槽底部添加消毒后的涂泥12 cm, 对虾养殖槽用气石连续增氧, 虾槽水位低于蛏槽水位形成一定的高度差, 利用水泵将虾槽的水泵入蛏槽, 蛏槽里的水自流回到虾槽, 实现虾槽和对应蛏槽形成水循环。实验装置系统如图 1所示。
 |
| 图 1 养殖单元装置示意图 Fig. 1 Schematic diagram of the breeding device 注: A为蛏槽; B为缓冲区; C为虾槽; D为支撑台; E为水泵; 箭头①代表水泵泵水方向; 箭头②代表水流方向 |
实验用海水是自然海水经两次过滤后使用, 每个养殖单元每天换水量40%~80%。换后废水采用净化技术加以处理, 采用多种作用有效去除废水中颗粒物及蓝绿藻等。实验期间水温24.0~28.1 ℃, pH 7.64~8.27, 盐度20, 虾槽溶解氧大于5.99 mg/L, 蛏槽溶解氧大于4.90 mg/L。
凡纳滨对虾投喂“强普”牌饲料, 日投喂量按照对虾体重的3%~4%进行投喂, 每天分3次投喂, 投喂时间分别为6:00、12:00、16:00。缢蛏投喂人工培养的角毛藻(Chaetoceros)和微绿球藻(Nannochloropsis gaditana), 用“宁波大学M3#”营养盐配方培养两种微藻, 藻密度维持在10×104 cell/mL, 投喂量依照天气、水质以及缢蛏摄食情况作适当调整。
1.2.2 水质样品采集与测定实验期间每天用便捷式溶氧仪测定温度和溶解氧; 盐度计、pH计测定盐度和pH。在实验开始的第1、7、14、21、28天取虾槽和蛏槽水样。依据海洋监测规范(GB 17378.4-2007)测定水样中TN、TP、氨氮、硝酸盐、亚硝酸盐、活性磷酸盐等水质因子。
1.2.3 虾蛏生长指标实验开始及结束时测定对虾和缢蛏, 将养殖虾蛏称重并记录, 计算增重率及特定生长率。
计算公式为:
 (1)
(1)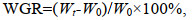 (2)
(2)式中, Wt: 实验结束时虾蛏重量; W0: 实验开始时虾蛏重量; t: 养殖天数。
1.2.4 免疫指标及消化酶活性测定实验结束时, 3个实验组中的每个平行组随机选取5个缢蛏的内脏团、5尾凡纳滨对虾的肝胰腺和肌肉组织, 迅速置于液氮中保存, 然后将所取样品放入–80 ℃超低温冰箱保存。采样过程遵循欧盟关于动物实验的第2010/63/EU号指令和国家研究委员会的实验动物护理和使用指南。所测免疫指标包括超氧化物歧化酶(SOD)、过氧化氢酶(CAT)、酸性磷酸酶(ACP)、碱性磷酸酶(AKP)、丙二醛(MDA)。所测的消化酶有胃蛋白酶、胰蛋白酶、脂肪酶(LPS)、淀粉酶(AMS)。所有用于酶活测定试剂均购自南京建成生物工程所, 各项指标都是按照说明书测定方法进行测定。
1.2.5 高通量测序样品本实验所用样品于养殖实验结束采集(表 2), 采集的样本包括了9个养殖单元的水体(SC1-1~SC3-3)、养殖单元中蛏槽泥样(N1-1~N3-3)和养殖单元中缢蛏内脏团样品(C1-1~ C3-3)共27个样品。采样时养殖单元水温(23.5±0.5) ℃, pH为8.11±0.19, 每个养殖单元的水样包含了虾槽水样和蛏槽水样, 水样和泥样采用五点法采集。所采的水样, 用0.22 μm微孔滤膜的真空抽滤装置进行抽滤, 然后将滤膜装进冻存管放入–80 ℃超低温冰箱待测。取表层1 cm底泥, 混匀后置于冻存管中, 放入–80 ℃超低温冰箱待测。缢蛏内脏团样本的采集即从每个养殖单元随机5只缢蛏, 在无菌状态下取出缢蛏内脏团, 并装入冻存管中。然后将样品送美格基因科技有限公司进行高通量测序。
| 样品 | 缩写 |
| 水体 | SC |
| 底泥 | N |
| 缢蛏内脏团 | C |
实验数据采用SPSS 26软件进行单因素方差分析(One-Way ANOVA), 结果以平均值±标准差(mean±SD)表示。采用邓肯假定等方差和邓尼特不假定等方差事后比较各实验组数据之间差异, P < 0.05为显著差异。
2 结果与分析 2.1 各实验组生长指标比较各实验组对虾和缢蛏的生长指标如表 3所示。从三个实验组对虾的生长看, LP组特定生长率和增重率最高, 且MP组与LP组无显著性差异(P > 0.05)。各实验组缢蛏的生长与对虾相似, LP组和MP组比较, 两组的特定生长率和增重率十分接近, HP组的特定生长率和增重率仅为0.60%和18.34%。无论是对虾还是缢蛏, 在对虾数量一定的情况下均表现出HP组的特定生长率与增重率都是最低, 即增加缢蛏数量会影响虾蛏的生长, 且与其他两组呈显著性差异(P < 0.05)。
| 组别 | 特定生长率/% | 增重率/% |
| LP-X | 1.29±0.26* | 43.88±10.68* |
| MP-X | 1.25±0.17* | 41.91±6.74* |
| HP-X | 0.61±0.08 | 18.80±2.16 |
| LP-C | 0.92±0.16* | 29.34±5.73* |
| MP-C | 0.92±0.20* | 29.40±7.26* |
| HP-C | 0.60±0.02 | 18.34±0.65 |
| 注: LP: 低虾蛏数量配比组; MP: 中虾蛏数量配比组; HP: 高虾蛏数量配比组; X: 凡纳滨对虾; C: 缢蛏; *: P < 0.05 | ||
三个虾蛏不同数量配比实验组的水质因子变化如图 2所示。各实验组总氮、总磷、硝酸盐、亚硝酸盐、氨氮和活性磷酸盐浓度前期和中期总体呈上升趋势, 后期的亚硝酸盐和硝酸盐出现明显下跌。组间水质营养盐浓度特点表现为HP组 > MP组 > LP组。氨氮浓度变化在0.05~1.06 mg/L, 第1~7 d时其浓度迅速升高, 第7 d后浓度保持相对稳定。亚硝酸盐和硝酸盐在第21 d时达到峰值, 第21 d后各浓度出现不同程度下降。总氮浓度表现为各实验组到达峰值时间不同, HP组总氮浓度在第14 d达到峰值, MP组第21 d达到峰值而LP组总氮浓度一直升高。活性磷酸盐和总磷浓度在养殖过程中一直升高, 第21 d后升高趋势变缓。
 |
| 图 2 虾蛏串养系统主要水质因子含量变化 Fig. 2 Changes in the content of the main water quality factors in the shrimp- razor-clam tandem culture system |
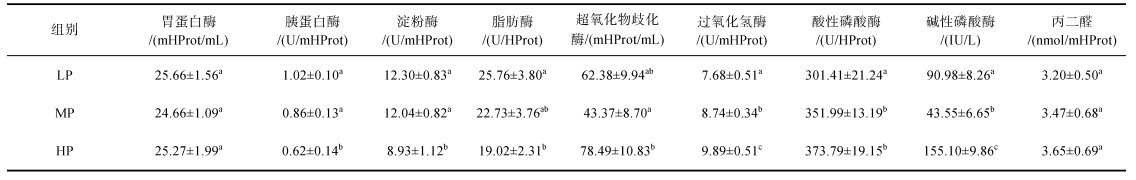
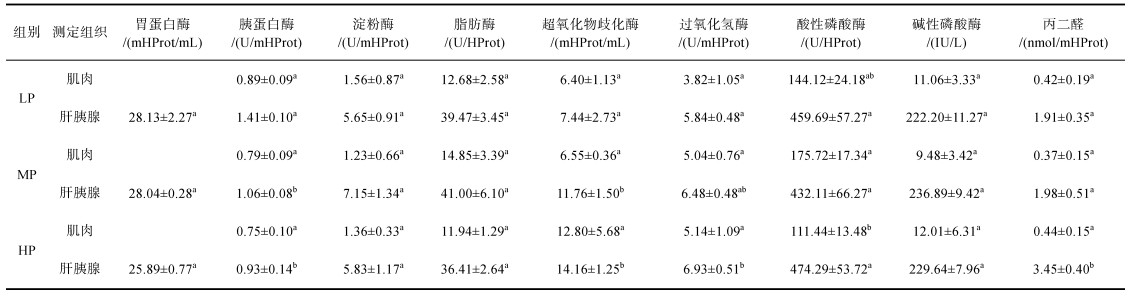
凡纳滨对虾和缢蛏的消化酶变化呈现不同的特点, 凡纳滨对虾和缢蛏有不同的方式来应对环境变化。四种消化酶中除胃蛋白酶无明显变化, 其余3种酶在各实验组缢蛏中均有显著性差异(P < 0.05)。胰蛋白酶在LP组缢蛏中活性最高, 显著高于MP组和HP组(P < 0.05)。淀粉酶在HP组活性最低, 与其余两组存在显著性差异(P < 0.05)。脂肪酶活性在LP组和HP组中存在显著性差异(P < 0.05)。不同实验组对虾肌肉中消化酶变化不大, 但在对虾肝胰腺中胰蛋白酶活性不同实验组间有着显著差异, 对虾肝胰腺中胰蛋白酶活性在HP组最低, 与其余两组呈显著差异(P < 0.05)。
由表 4和表 5可知, 虾蛏在不同实验组中非特异性免疫表现方式也不同。与对虾肌肉比较而言, 缢蛏内脏团和对虾肝胰腺中非特异性免疫酶对环境变化更为敏感。缢蛏内脏团中除MDA无明显变化外, CAT和AKP在不同实验组中存在显著性差异(P < 0.05), SOD在MP组与HP组存在显著性差异(P < 0.05), ACP在LP组中活性最低, 且与其余两组存在显著性差异(P < 0.05)。对虾肌肉非特异性免疫酶酶活在不同实验组中表达较稳定, 仅HP组中的ACP与其余两组存在显著性差异(P < 0.05)。ACP和AKP在不同实验组对虾肝胰腺中表达稳定, 各组间不存在显著性差异(P > 0.05)。SOD和MDA在HP组中活性最高, 与另两组存在显著性差异(P < 0.05)。CAT在LP组和HP组中呈显著性差异(P < 0.05)。
|
|
由图 3可知, 各实验组共有197个OTUs, 水样、底泥和缢蛏内脏团微生物OTU数量特征趋势一致。随着虾蛏数量配比的增加, OTU数量逐渐增加。水中richness指数在MP组与其余两组存在显著性差异(P < 0.05)。底泥和缢蛏内脏团所有处理(chao1、simpson、richness、reads)之间无显著性差异(P > 0.05)。
 |
| 图 3 水样、底泥和缢蛏内脏团微生物OTU比较 Fig. 3 Comparison of microbial OTUs in water, substrate, and razor clam visceral mass 注: SC: 水样; N: 底泥样; C: 缢蛏内脏团样 |
水样、底泥、缢蛏内脏团样品在门水平上的微生物群落丰度及组成见图 4所示。对水样、底泥、缢蛏内脏团3种样品的高通量测序数据进行分析, 所有序列共分为57门121纲242目367科628属8 026个OTUs。由图 4显示, 水、底泥及缢蛏内脏团微生物相对丰度不尽相同, 样品间优势菌门不同。随着虾蛏数量配比增加, 缢蛏内脏团中软壁菌门不再具有优势地位, 而变形菌门、螺旋体门、Epsilonbacteraeota丰度仍占优势地位(P > 0.05), 且放线菌门和厚壁菌门丰度有所增加。底泥微生物种类多但丰度相差大, 变形菌门和拟杆菌门占80%以上, 其余微生物丰度占比不足20%。水中微生物种类较少, 变形菌门分度占比70%以上, 分别达到80.0%、73.9%和84.6%。而不同实验组水样中拟杆菌门相对丰度相差不大, 分别占12.5%、12.8%和11.1%, 表明水环境因子对拟杆菌门无明显影响。
 |
| 图 4 水样、底泥、缢蛏内脏团样品在门水平上的微生物群落丰度及组成 Fig. 4 Microbial community abundance and composition at the phylum level for water, substrate, and razor clam visceral mass samples |
水样、底泥、缢蛏内脏团样品中门水平上微生物群落丰度的聚类如图 5所示。从水样、底泥、缢蛏内脏团三种样品中选取丰度前15的物种进行聚类分析。首先, 在样品聚类方面, 水样和底泥之间的相似性与缢蛏相比较而言更加紧密, 也就是水和底泥之间的影响更大。在缢蛏样品方面, MP与HP组聚类之后, 再和LP组聚类。这种情况同样在底泥样品中出现, 水样聚类略有差别, LP组与HP组聚类后和MP组聚类。水、底泥和缢蛏内脏团样品微生物群落比较看出, 缢蛏内脏团微生物丰度高的微生物类群较多, 包含了厚壁菌门、放线菌门、Epslionbacteraeota、软壁菌门、蓝细菌门、螺旋体门、绿弯菌门及浮霉菌门等。LP组中软壁菌门相对丰度高于其余两组底泥中微生物丰度高的微生物类群数量和缢蛏内脏团微生物丰度高的微生物类群相当, 绿弯菌门、浮霉菌门、疣微菌门、放线菌门、Dependentiae、拟杆菌门及Patescibacteria。水中微生物群落丰度相对较高的仅2门, 即变形菌门和Epslionbacteraeota。
 |
| 图 5 虾蛏不同数量配比各样品门水平上微生物群落丰度的聚类热图 Fig. 5 Cluster heat map of microbial community abundance at phylum level of each sample in different quantitative ratios of shrimp and razor clam 注: Firmicutes: 厚壁菌门; Actinobacteria: 放线菌门; Tenericutes: 软壁菌门; Cyanobacteria: 蓝细菌门; Spirochaetes: 螺旋体门; Proteobacteria: 变形菌门; Chloroflexi: 绿弯菌门; Planctomycetes: 浮霉菌门; Verrucomicrobia: 疣微菌门; Acidobacteria: 酸杆菌门; Bacteroidetes: 拟杆菌门 |
水样、底泥及缢蛏内脏团样品在属水平上的微生物群落丰度及组成如图 6所示。在属水平上, 各实验组样品中微生物丰度大于1%的优势细菌组成及丰度存在明显不同。弓形菌属作为三个实验组缢蛏内脏团优势菌属, 实验组之间无显著差异(P > 0.05)。支原体属丰度随着虾蛏数量配比增加而呈降低趋势, Draconibacterium和罗姆布茨菌属变化情况与支原体属情况相反。底泥中Tenacibaculum和黄杆菌属丰度降低表现在MP组和HP组中, 其余丰度占比较稳定。水样中优势菌属是Donghicola, 另外水样中海杆菌属丰度随着虾蛏数量配比增加而增加。Marivita也属于水样微生物的一种优势菌, LP组丰度低于其余两组。
 |
| 图 6 水样、底泥及缢蛏内脏团样品在属水平上的微生物群落丰度及组成 Fig. 6 Microbial community abundance and composition at the genus level in water, substrate, and razor clam viscera mass 注: Arcobacter: 弓形菌属; Mycoplasma: 支原体属; Marinobacterium: 海杆菌属; Vibrio: 弧菌属; Polaribcter_4: 极地杆菌属; Flavobacterium: 黄杆菌属; Romboutsia: 罗姆布茨菌属; Cyanobium_PCC-6307: 蓝细菌属 |
水质环境因子与优势菌群相关性分析见图 7。由图 7可知, 厚壁菌门、绿弯菌门、Dependentiae、拟杆菌门、酸杆菌门、Patescibacteria及浮霉菌门与TN、TP正相关性高(P < 0.05), 其中厚壁菌门、绿弯菌门、Patescibacteria和浮霉菌门还与NO3—、PO4—、NO2—、NH4+呈正相关, Proteobacteria与水质环境因子呈正相关(P < 0.05), 但正相关性要低一些。Epsilonbacteraeota、软壁菌门、蓝细菌门与TN、TP、NO3—、PO4—、NO2—、NH4+均呈负相关(P < 0.05)。疣微菌门与TN、TP、NO3—呈正相关(P < 0.05), 而与PO4—、NO2—、NH4+呈负相关(P < 0.05)。螺旋体门与NO3—呈负相关, 与其余五项水质环境因子呈正相关。放线菌门与TN、TP呈负相关, 与NO3—、PO4—、NO2—、NH4+呈正相关(P < 0.05)。拟杆菌门和酸杆菌门与NO3—、PO4—、NO2—、NH4+相关性情况与放线菌门相反。Dependentiae与NO3—、PO4—呈正相关(P < 0.05), 而与NO2—、NH4+呈负相关(P < 0.05)。表明不同菌群对水质因子有着不一样的影响。
 |
| 图 7 水质环境因子与优势菌群相关性分析 Fig. 7 Correlation analysis between water-quality environmental factors and dominant bacterial population 注: Verrucomicrobia: 疣微菌门; Tenericutes: 软壁菌门; Cyanobacteria: 蓝细菌门; Spirochaetes: 螺旋体门; Actinobacteria: 放线菌门; Firmicutes: 厚壁菌门; Chloroflexi: 绿弯菌门; Bacteroidetes: 拟杆菌门; Acidobacteria: 酸杆菌门; Planctomycetes: 浮霉菌门 |
不同样品微生物与环境因子之间的RDA分析见图 8所示。图中是本研究27个样品中微生物与环境因子之间的关系, 得到影响样品分布的重要环境驱动因子。不同样品微生物与环境因子之间RDA分析显示, 横轴能解释物种变量为63.1%; 纵轴能解释物种变量为14%。水体中的TN、NO3—、PO4—对不同样品微生物影响更大。NO3—、PO4—、NO2—、NH4+与水样样品中微生物呈正相关, MP组水样样品微生物与TN、TP在底泥样品中, 各组底泥样品与水环境因子相关性相似, 除HP组的一组平行样之外, 其余底泥样与TP、TN呈正相关, 与NO3—、PO4—、NO2—、NH4+均呈负相关。
 |
| 图 8 不同样品微生物与环境因子之间的RDA分析 Fig. 8 Correlation analysis between microorganisms and environmental factors of different samples 注: SC: 水样; N: 底泥样; C: 缢蛏内脏团样; 1表示LP组; 2表示MP组; 3表示HP组 |
虽然本研究的虾蛏串联养殖效果已经在生产实践中得到肯定, 充分利用了水体的空间, 相比于其他传统养殖模式提高了水体利用率。对虾养殖塘既给对虾生长提供空间又使藻类能充分利用对虾残饵粪便作为氮磷营养源, 继而供蛏塘中缢蛏滤食, 减少了养殖废水的排放, 实现绿色健康养殖目标(骆云慧等, 2021)。符合未来水产养殖可持续发展的方向。那么, 在这样的串联养殖模式中对虾与缢蛏合理的数量配比是多少?也就是说, 在对虾密度不变的情况下, 最适的缢蛏密度是多少?这正是本实验要解决的科学问题。在本实验中, 低数量配比的LP组生长指标更好, MP组与LP组生长指标相差不大。LP组虾蛏特定生长率和增重率最高, 与高数量配比的HP组虾蛏特定生长率和增重率呈显著性差异(P < 0.05)。有研究认为高密度养殖会增加个体之间对资源以及空间的竞争, 造成较多的能量消耗而影响生长(Da Costa et al, 2016)。实验中HP组虾蛏生长受到明显抑制。其他研究也表明, 虾夷扇贝(Mizuhopecten yessoensis)在适宜养殖密度下耗氧率和排氨率较低(车建锋等, 2020)。何苗(2017)研究温度对缢蛏排氨率不存在着显著性差异(P > 0.05)。也就是说在本实验中, 排除了水温的变化对排氨率影响。在本实验的水质指标反映, 高虾蛏数量配比HP组中, 因缢蛏数量增加, 导致养蛏水体中氨氮含量高于其余两组, 由于虾蛏串养模式属于循环水养殖, 导致整个养殖单元水体中氨氮含量同步升高。但氨氮在养殖第21天后浓度下降的原因来自于养殖换水、贝类的吸收、环境微生物降解以及底泥吸附。顾楠(2020)研究氨氮对凡纳滨对虾生长及存活有着明显影响。当甲壳动物受到氨氮胁迫时, 甲壳动物体内氨氮解毒代谢途径会发生一些变化。该途径会降低甲壳动物鳃组织对分子氨通透性, 增加NH4+排除率(岳峰, 2010)。也就表明HP组水环境比LP、MP组水环境差, 虾蛏生长不仅受到密度胁迫, 而且对虾也受到氨氮胁迫, 最终造成实验结束时HP组对虾和缢蛏的生长均不如LP组和MP组。同时, 滤食性贝类对养殖生态修复有着积极的作用, 李黎等(2015)的研究表明, 适宜密度的放养缢蛏(Sinonovacula constricta)能较好促进沉积物中磷的循环和再生, 提高水体氮的利用率, 有效调控水体中亚硝酸盐及氨氮等有害物质含量。方磊等(2011)研究了河蚌(Anodonta woodiana)对中华鲟养殖水体生态修复效果, 发现背角无齿蚌能显著降低水体总磷的含量。另外, 与传统虾蛏混养模式相比, 虾蛏串联养殖模式更加有利于维持池塘生态稳定, 对养殖池塘水生态环境有明显修复作用(鲍格格等, 2022)。从本研究水质指标来看, HP组超出系统中缢蛏对氨氮和无机磷的调控, 严重影响虾蛏生长。因此, 本实验结果表明, 从兼顾虾蛏生长与养殖产量考虑, 在对虾养殖密度60 ind./m2条件下, 虾蛏数量配比以MP组1︰5最为合适, 即缢蛏密度300 ind./m2。
3.2 从消化酶活和非特异性免疫指标评价虾蛏数量配比消化酶和非特异性免疫酶活性在一定程度上反映了养殖对象健康状况。当养殖对象受到一定程度的环境胁迫时, 消化酶及非特异性免疫酶活性变化也会有所表现。如有研究发现氨氮胁迫使得黄颡鱼(Pelteobagrus fulvidraco)幼鱼肠道消化酶活性破坏(Wang et al, 2021)。在氨氮胁迫对方斑东风螺消化酶的影响研究中显示, 随着低质量氨氮浓度(22 mg/L)胁迫时间的延长, 方斑东风螺胃蛋白酶活性总体上呈现抑制作用(谭春明等, 2019)。王贞杰等(2017)研究表明, 氨氮胁迫会使圆斑星鲽(Verasper variegatus)幼鱼肝脏中的MDA含量显著升高。从对虾与缢蛏的消化酶活性分析发现, 虾蛏可能是受到生存空间竞争以及食物竞争的影响, 导致不能达到较好状态。有研究认为, 高密度会显著降低凡纳滨对虾生长和免疫性能(Liu et al, 2017)。本研究中各实验组之间, 缢蛏内脏团和凡纳滨对虾中SOD和CAT存在显著差异, HP组SOD及CAT活性显著低于MP和LP组(P < 0.05)。表明不同虾蛏数量配比也同样影响着缢蛏和凡纳滨对虾消化酶及非特异性免疫酶活性。活性氧攻击生物细胞膜中多不饱和脂肪酸的时候, 会引起脂质的过氧化, 从而形成一些脂质过氧化物, 比如醛基(丙二醛)、酮基、羟基、羰基、氢过氧基等(马细兰等, 2018)。SOD是已知的唯一一种可以利用自由基作为底物并催化超氧阴离子歧化的酶, 其对机体的氧化和抗氧化平衡起着至关重要的作用, 能清除超氧阴离子自由基, 保护细胞免受损伤(Li et al, 2012)。CAT和GSH协同催化H2O2分解到H2O和O2 (Xi et al, 2012)。在活生物体内自由基与脂质反应产生脂质过氧化, 氧化的最终产物是MDA, 它引起大分子的交联聚合, 如蛋白质和核酸, 并诱导细胞毒性(Heinlaan et al, 2008)。同样, MDA作为一种最重要的脂质过氧化产物, 是油脂中多元不饱和脂肪酸过氧化产物, 也可以反映机体脂质的过氧化水平高低(Lepage et al, 1991)。表明高虾蛏数量配比组受较大的环境压力, 胁迫可导致虾蛏体内生理活动受到干扰, 甚至受到了损伤。因此, 从消化酶以及免疫酶指标综合来看, MP组虾蛏有着较好的摄食以及非特异性免疫状态, 说明更适应对虾养殖密度60 ind./m2和缢蛏密度300 ind./m2的串联养殖环境。
3.3 对虾与缢蛏数量配比与微生物组成特点的关系本研究结果表明, 三个不同虾蛏数量配比实验组中, 水样、底泥和缢蛏内脏团微生物丰度及组成之间存在着差异。水样和底泥之间的关系更为密切, 两者的微生物优势菌门相同, 均为变形菌门(Proteobacteria)和拟杆菌门(Bacteroidetes), 两者占各自微生物总数80%以上。据报道, 变形菌是沉积物或土壤中最丰富的门类, 因为它可以适应不同的极端环境, 并强烈参与沉积物中有机物降解和代谢过程(Dong et al, 2019)。在养殖实验后期虾蛏串养系统中菌群较为稳定, 水质营养盐受到菌群作用出现不同程度的下降。比如海杆菌属是属于反硝化细菌, 能将硝酸盐中的氮通过一系列中间产物还原为氮气, 长时间能降低水体中总氮(巫亮, 2014)。在属水平上, 水样和底泥微生物组成差异比较明显, 底泥中微生物多样性高于水样和缢蛏内脏团微生物。主要是底泥中物质组成较复杂, 营养较为丰富, 比较适合微生物生长, 提高了底泥微生物组成丰度及种类, 对分解利用底泥中沉积的有机物起到积极的作用。虽然不同数量配比实验组中缢蛏内脏团优势菌丰度也表现不同, 但缢蛏内脏团微生物优势菌门都有变形菌门(Proteobacteria)、软壁菌门(Tenricutes)、螺旋体门(Spirochaetes)。陈琼等(2017)发现的虾养殖池水体的微生物物种相对丰度前10包含有疣微菌门和软壁菌门。另外, 刘文亮等(2017)在凡纳滨对虾肠道中发现疣微菌门, 表明这两种菌与虾联系紧密。从图 3看出, 串养系统水样以及缢蛏内脏团中也发现了疣微菌门的存在, 表明缢蛏内脏团微生物组成变化与虾蛏串养有关, 微生物组成也会受到养殖对象的改变而改变。本研究表明, 微生物组成也会受到养殖密度的影响, 在不同实验组中, 缢蛏内脏团微生物丰度中有所变化。并发现随着虾蛏数量配比的增加, 导致软壁菌门丰度逐渐降低, 同时水体中TN出现不同程度的升高, 说明对氮磷循环产生一定的影响。有研究指出, 软壁菌门具有突出的核酸降解能力, 对磷、氮等元素的循环有着重要驱动作用(Zheng et al, 2021)。厚壁菌门、放线菌门丰度可能与虾蛏数量配比呈正相关, 厚壁菌门能产生芽孢, 可抵抗脱水和极端环境, 有固氮作用。因此也可看出某些微生物在水质调控方面起着积极作用。本研究中弓形菌丰度的增加可能是有硫化物产生。弓形菌属的代谢多功能性可能在以高硫化物、低氧和动态流体运动为特征的水体中提供竞争优势(Jurado et al, 2021), 也较为符合本研究系统循环水特征。由于HP组比其他组获得的更多的食物来源, 使得有机物变得丰富, 放线菌主要功能是吸收营养物质, 使得放线菌在较高密度组中更容易生长。不同实验组水样中海杆菌属丰度也有所不同, 与TN呈正相关。董学兴等(2019)研究发现, TP显著影响罗氏沼虾肠道微生物结构, 同时养殖模式可通过影响水体营养盐尤其是氮磷的含量影响罗氏沼虾肠道微生物群落结构, 与本研究结果类似, 相关性分析证实TN是水样、底泥和缢蛏内脏团微生物群落结构改变的主要水质因子, 氮磷元素水平间接地影响着环境及肠道微生物菌群的构成。总之, 水、底泥和缢蛏内脏团存在一定关系, 水和底泥微生物相似性更高, 联系更加紧密。底泥微生物组成及种类更加丰富, 很大可能影响着水中和缢蛏的微生物组成。在虾蛏数量配比下影响微生物的主要驱动因子是总氮和活性磷酸盐。MP组虾蛏数量配比为1︰5 (60 ind./m2︰300 ind./m2)时, 串联养殖系统中微生物群落组成对环境更友好。
4 结论综上所述, 不同虾蛏数量配比使得水质营养盐浓度发生不同程度变化, 进而使虾蛏消化酶和非特异性免疫酶活性升高或降低, 改变虾蛏及环境微生物组成及丰度, 在养殖后期, 稳定的菌群对水质营养盐产生反馈调节, 使得水质营养盐浓度有降低的趋势, 减少虾蛏来自环境的压力。受到上述多种因素影响, 在虾蛏增重率和特定生长率方面表现明显。在虾蛏数量配比串联养殖模式中, 低中虾蛏数量配比组特定生长率、增重率等生长指标明显高于高虾蛏数量配比组。从养殖环境内外因素分析, 低中虾蛏数量配比串联养殖模式更优。采用低中虾蛏数量配比(60 ind./m2 : 180 ind./m2; 60 ind./m2 : 300 ind./m2)的虾蛏串联养殖模式, 能维持较高的虾蛏生长水平、有效控制营养盐因子, 减少对虾蛏应激损伤, 给虾蛏营造良好的养殖环境。
马细兰, 郑陆威, 叶超霞, 等, 2018. 胃促生长素Ghrelin在不同盐度胁迫下对黑鲷(Acanthopagrus schlegelii)部分免疫指标的影响. 海洋与湖沼, 49(3): 692-699 |
王贞杰, 陈四清, 曹栋正, 等, 2017. 急性氨氮胁迫对圆斑星鲽(Verasper variegatus)幼鱼鳃和肝组织结构及相关酶活性的影响. 渔业科学进展, 38(2): 59-69 |
王志滨, 许鹏, 郑冬梅, 等, 2014. 微藻在几种常见贝类人工育苗中的应用. 现代农业科技, (11): 299-300 |
车建锋, 王晓辉, 肖露阳, 等, 2020. 不同养殖密度对虾夷扇贝生长、耗氧率和排氨率的影响. 江苏农业科学, 48(17): 179-183 |
方磊, 刘健, 陈锦辉, 等, 2011. 中华鲟养殖池背角无齿蚌和鲢鳙鱼生态修复效果比较. 生态科学, 30(3): 288-294 DOI:10.3969/j.issn.1008-8873.2011.03.012 |
任丽娟, 何聃, 邢鹏, 等, 2013. 湖泊水体细菌多样性及其生态功能研究进展. 生物多样性, 21(4): 422-433 |
刘文亮, 许华, 唐杨, 等, 2017. 饲料中补充蜡样芽孢杆菌(Bacillus cereus)生物膜对凡纳滨对虾(Litopenaeus vannamei)生长、抗病力及其肠道微生物组成的影响. 渔业科学进展, 38(4): 87-95 |
刘招坤, 2014. 独立供饵式缢蛏池塘蓄水养殖系统设计与试验研究. 渔业现代化, 41(6): 18-21 DOI:10.3969/j.issn.1007-9580.2014.06.004 |
巫亮, 2014. 海杆菌属反硝化能力及其酶系的分布特点的研究[D]. 南京: 南京工业大学: 4-5.
|
李黎, 刘楠, 李东明, 等, 2015. 养殖密度对缢蛏养殖系统水质的影响. 生态学杂志, 34(10): 2893-2899 |
何苗, 2017. pH、温度、饵料浓度对缢蛏钙化率及能量代谢的影响[D]. 上海: 上海海洋大学: 20-21.
|
张清科, 金珊, 王建平, 等, 2019. 供饵式缢蛏养殖试验. 水产养殖, 40(9): 14-15 DOI:10.3969/j.issn.1004-2091.2019.09.007 |
陈志, 2017. 盐度、密度和底质对双线紫蛤幼贝存活及生长的影响. 渔业研究, 39(1): 34-39 |
陈琼, 李贵阳, 罗坤, 等, 2017. 凡纳滨对虾(Litopenaeus vannamei)亲虾繁殖期水体微生物多样性. 海洋与湖沼, 48(1): 130-138 |
岳峰, 2010. 三疣梭子蟹在氨氮胁迫下免疫应答与解毒代谢机制的研究[D]. 青岛: 中国海洋大学: 10-11.
|
金若晨, 2020. 凡纳滨对虾养殖池塘水质及养殖环境和虾肠道微生物群落结构研究[D]. 上海: 上海海洋大学: 10-12.
|
骆云慧, 吴波, 徐善良, 等, 2021. 缢蛏动态能量收支生长模型的建立. 水产学报, 45(4): 578-587 |
顾楠, 2020. 氨氮胁迫对凡纳滨对虾生长、生理指标、鳃和肝胰腺显微结构和相关基因表达的影响[D]. 上海: 上海海洋大学: 16-17.
|
曹豪豪, 张红兵, 薛溪发, 等, 2021. 新型基因编辑技术在单细胞微藻中的应用进展. 生物技术进展, 11(1): 9-15 |
董学兴, 吕林兰, 赵卫红, 等, 2019. 不同养殖模式下罗氏沼虾肠道菌群结构特征及其与环境因子的关系. 上海海洋大学学报, 28(4): 501-510 |
鲍格格, 赵淳朴, 梅泊承, 等, 2022. 基于权重赋值法与灰色聚类法评价虾蛏串联养殖池塘的生态修复效果. 海洋与湖沼, 53(1): 206-214 |
谭春明, 赵旺, 于刚, 等, 2019. 氨氮胁迫对方斑东风螺溶菌酶及3种常见消化酶活力的影响. 南方水产科学, 15(3): 120-125 |
薛宝宝, 2019. 缢蛏新品种"申浙1号"性状分析及耐高温相关基因SNP位点筛选[D]. 上海: 上海海洋大学: 2-3.
|
BROWN T W, CHAPPELL J A, BOYD C E, 2011. A commercialscale, in-pond raceway system for Ictalurid catfish production. Aquacultural Engineering, 44(3): 72-79 |
CHEN F F, LENG Y Y, LU Q, et al, 2021. The application of microalgae biomass and bio-products as aquafeed for aquaculture. Algal Research, 60: 102541 |
CROVESY L, MASTERSON D, ROSADO E L, 2020. Profile of the gut microbiota of adults with obesity: a systematic review. European Journal of Clinical Nutrition, 74(9): 1251-1262 |
DA COSTA F P, DE FARIAS GOMES B S F, DO NASCIMENTO ALVES PEREIRA S D, et al, 2016. Influence of stocking density on the behaviour of juvenile Litopenaeus vannamei (Boone, 1931). Aquaculture Research, 47(3): 912-924 |
DAI W F, DONG Y H, YE J, et al, 2022. Gut microbiome composition likely affects the growth of razor clam Sinonovacula constricta. Aquaculture, 550: 737847 |
DONG X Y, GREENING C, RATTRAY J E, et al, 2019. Metabolic potential of uncultured bacteria and archaea associated with petroleum seepage in deep-sea sediments. Nature Communications, 10(1): 1816 |
DRAPCHO C M, BRUNE D E, 2000. The partitioned aquaculture system: impact of design and environmental parameters on algal productivity and photosynthetic oxygen production. Aquacultural Engineering, 21(3): 151-168 |
GUPTA S, PAWAR S B, PANDEY R A, 2019. Current practices and challenges in using microalgae for treatment of nutrient rich wastewater from agro-based industries. Science of the Total Environment, 687: 1107-1126 |
HEINLAAN M, IVASK A, BLINOVA I, et al, 2008. Toxicity of nanosized and bulk ZnO, CuO and TiO2 to bacteria Vibrio fischeri and crustaceans Daphnia magna and Thamnocephalus platyurus. Chemosphere, 71(7): 1308-1316 |
HUCHETTE S M H, KOH C S, DAY R W, 2003. Growth of juvenile blacklip abalone (Haliotis rubra) in aquaculture tanks: effects of density and ammonia. Aquaculture, 219(1/2/3/4): 457-470 |
JURADO V, D'ANGELI I, MARTIN-POZAS T, et al, 2021. Dominance of Arcobacter in the white filaments from the thermal sulfidic spring of Fetida Cave (Apulia, southern Italy). Science of the Total Environment, 800: 149465 |
LEPAGE G, MUNOZ G, CHAMPAGNE J, et al, 1991. Preparative steps necessary for the accurate measurement of malondialdehyde by high-performance liquid chromatography. Analytical Biochemistry, 197(2): 277-283 |
LI Y F, GU Z Q, LIU H, et al, 2012. Biochemical response of the mussel Mytilus coruscus (Mytiloida: Mytilidae) exposed to in vivo sub-lethal copper concentrations. Chinese Journal of Oceanology and Limnology, 30(5): 738-745 |
LIU G, ZHU S M, LIU D Z, et al, 2017. Effects of stocking density of the white shrimp Litopenaeus vannamei (Boone) on immunities, antioxidant status, and resistance against Vibrio harveyi in a biofloc system. Fish & Shellfish Immunology, 67: 19-26 |
NADELLA R K, PANDA S K, RAO B M, et al, 2021. Antibiotic resistance of culturable heterotrophic bacteria isolated from shrimp (Penaeus vannamei) aquaculture ponds. Marine Pollution Bulletin, 172: 112887 |
NEWTON R J, JONES S E, EILER A, et al, 2011. A guide to the natural history of freshwater lake bacteria. Microbiology and Molecular Biology Reviews, 75(1): 14-49 |
SAHIN K, YAZLAK H, ORHAN C, et al, 2014. The effect of lycopene on antioxidant status in rainbow trout (Oncorhynchus mykiss) reared under high stocking density. Aquaculture, 418/419: 132-138 |
SHALABY E, 2011. Algae as promising organisms for environment and health. Plant Signaling & Behavior, 6(9): 1338-1350 |
SMITH D W, PIEDRAHITA R H, 1988. The relation between phytoplankton and dissolved oxygen in fish ponds. Aquaculture, 68(3): 249-265 |
TURKER H, EVERSOLE A G, BRUNE D E, 2003. Filtration of green algae and cyanobacteria by Nile tilapia, Oreochromis niloticus, in the Partitioned Aquaculture System. Aquaculture, 215(1/2/3/4): 93-101 |
WANG S D, LI X, ZHANG M Z, et al, 2021. Ammonia stress disrupts intestinal microbial community and amino acid metabolism of juvenile yellow catfish (Pelteobagrus fulvidraco). Ecotoxicology and Environmental Safety, 227: 112932 |
XI S H, LIU Z Y, YAN L, et al, 2012. A role of fluoride on free radical generation and oxidative stress in BV-2 microglia cells. Mediators of Inflammation, 2012: 102954 |
YUAN Y, LUO J X, ZHU T T, et al, 2020. Alteration of growth performance, meat quality, antioxidant and immune capacity of juvenile Litopenaeus vannamei in response to different dietary dosage forms of zinc: Comparative advantages of zinc amino acid complex. Aquaculture, 522: 735120 |
ZHENG R K, LIU R, SHAN Y Q, et al, 2021. Characterization of the first cultured free-living representative of Candidatus izemoplasma uncovers its unique biology. The ISME Journal, 15(9): 2676-2691 |
ZHENG J P, ZHANG J, GUO Y L, et al, 2020. Improvement on metabolic syndrome in high fat diet-induced obese mice through modulation of gut microbiota by Sangguayin decoction. Journal of Ethnopharmacology, 246: 112225 |