 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 宋晨, 王晓波, 张国豪, 韩庆喜. 2023.
- SONG Chen, WANG Xiao-Bo, ZAHNG Guo-Hao, HAN Qing-Xi. 2023.
- 浙江瓯江口海域游泳动物群落特征及其与环境因子的关系
- COMMUNITY CHARACTERISTICS OF NEKTON ASSEMBLAGES IN OUJIANG RIVER ESTUARY AND ITS RELATIONSHIP WITH ENVIRONMENTAL FACTORS, ZHEJIANG, EAST CHINA
- 海洋与湖沼, 54(4): 1113-1124
- Oceanologia et Limnologia Sinica, 54(4): 1113-1124.
- http://dx.doi.org/10.11693/hyhz20221000258
文章历史
-
收稿日期:2022-10-11
收修改稿日期:2022-12-30




2. 禹治环境科技(浙江)有限公司 浙江宁波 315199;
3. 青岛道万科技有限公司 山东青岛 266237
2. Yuzhi Environmental Technology (Zhejiang) Co., Ltd., Ningbo 315199, China;
3. Qingdao Daowan Technology Co., Ltd., Qingdao 266237, China
瓯江口及邻近海域位于浙江省温州湾北部, 北接乐清湾, 东有洞头列岛和大北列岛为屏障, 水域开阔, 水深大多在20 m以内。瓯江口外侧潮汐为正规半日潮, 为强潮海湾, 平均潮差3.99~4.52 m, 最大潮差达7.21 m (龙湾)。潮流为不正规半日浅海潮流, 平均流速0.37~0.92 m/s (中国海湾志编纂委员会, 1993)。瓯江年均径流量202.7×108 m3, 带来了丰富的营养盐, 使得该海域水质肥沃; 潮流受江浙沿岸流和台湾暖流的共同制约, 在近岸地区形成上升流, 浮游生物丰富, 生物多样性水平和生产力较高(徐兆礼, 2008), 是海洋无脊椎动物和鱼类栖息、索饵、产卵、生长的优良场所, 海洋水产资源丰富, 经济水产动物种类繁多, 包括大黄鱼、小黄鱼、墨鱼、三疣梭子蟹、锯缘青蟹、缢蛏、海蜇等, 在瓯江口附近海域形成了792 km2的瓯江口渔场, 又称洞头渔场, 是东海第二大渔场(温州渔场)的重要组成部分(徐兆礼, 2008; 刘志坚等, 2016)。但是, 瓯江口附近海域在水产资源开发利用方面存在较多问题, 在长期高强度拖网捕捞(樊伟等, 2003)、沿海工业和滩涂围垦开发等人类活动影响下(徐兆礼, 2008), 生态系统已遭到破坏, 处于亚健康或不健康状态(刘志坚等, 2016), 海洋生物资源破坏严重, 大黄鱼、小黄鱼、曼氏无针乌贼、带鱼和海蜇等传统经济水生动物已经数量稀少, 无法形成渔汛, 取而代之的是低龄化、低值化、小型化的鱼类(中国海湾志编纂委员会, 1993)。
近年来, 瓯江口海域游泳动物或海洋生物资源的研究仍旧比较缺乏, 仅有的瓯江口游泳动物的研究中, 要么研究对象仅涵盖游泳动物中的单一类群(徐兆礼, 2008, 2009; 徐兆礼等, 2009; 陈伟峰等, 2021), 要么瓯江口仅作为其研究海域的一小部分且瓯江口附近站位数量稀少(李超男等, 2017; 陈伟峰等, 2021)或者研究区域位于瓯江内(刘志坚等, 2016), 缺少有针对性的瓯江口海域游泳动物群落生态学的研究。瓯江口海域生态环境和渔业资源变动的因素比较复杂, 相关管理部门为恢复渔业资源在瓯江口海域进行积极的增殖放流活动(姜存楷, 1994), 仅2019年即增殖放流曼氏无针乌贼、大黄鱼、黑鲷、日本对虾等8.1亿尾, 因此, 只有最新的调查研究才能及时准确地掌握瓯江口游泳动物群落的最新状况。基于此目的, 我们在2020年秋季和2021年春季在瓯江口海域进行了渔业资源调查, 力求如实反映瓯江口海域渔业生物的种类组成及多样性, 以期为研究河口水域环境变化和增殖放流活动影响下的渔业资源变迁特征提供数据基础, 为管理部门渔业生物资源保护政策的制定提供科学依据。
1 材料与方法 1.1 调查区域和时间于2020年11月(秋季)和2021年3月(春季)对瓯江口海域的游泳动物进行了单拖网采样, 调查范围为瓯江口附近海域, 共设计相同拖网调查站位12个, 分布于东起瓯江口, 西至洞头列岛, 北自乐清湾, 南达南麂列岛, 即12.85°~121.13°E和27.75°~28.07°N之间的广阔海域(图 1)。
 |
| 图 1 2020~2021年瓯江口海域游泳动物调查站位分布图 Fig. 1 Sampling locations of nekton assemblages in Oujiang River estuary during 2020~2021 |
在2020年秋季和2021年春季租用“浙洞渔83006”, 使用单拖网[10.0 m (宽)×5.0 m (高)], 网目范围2~7 cm, 其中网囊网目为2 cm, 拖网长度20 m, 网板规格1.2 m×0.5 m, 拖曳作业时网口水平扩展宽度约为6 m, 每网拖曳0.5~1.0 h。海上底拖网调查过程中, 规范记录各站位网产量、拖网时间等, 调查按《海洋调查规范》(GB/T 12673-2007)和《海洋渔业资源调查规范》(SC/T 9403-2012)进行。调查渔获物总质量在40 kg以下时, 全部取样分析; 大于40 kg时, 从中挑出大型和稀有的标本后, 从渔获物中随机取出渔获物分析样本20 kg左右。对渔获物均采用冰鲜冷藏, 带回实验室对渔获物进行种类、渔获重量和尾数统计, 记录网产量, 并对每个种类进行主要生物学测定(体长、体重、成幼体等)。鱼类的最新拉丁名及归属参照WoRMS (http://www.marinespecies.org)。依据调查海域物种分布和经济种类等情况, 本次调查海域渔获物主要分为鱼类、甲壳动物、头足类和其他等4大类群进行分别描述, 其中, 甲壳动物又分为虾类、蟹类和口足类, 尽管口足目和十足目中的虾类在分类上相差较大, 但它们在栖息生境、营养级和资源数量变化上具有一定的相似性, 因此将口足目和十足目虾类甲壳动物合并为“游泳虾形类”(简称虾类)(徐兆礼等, 2009)。其他类包括贝类、棘皮动物、星虫类等底栖无脊椎动物。
在采样的同时对站位的物理化学因子进行测量, 有机碳采用重铬酸钾氧化-还原容量法; 硫化物采用亚甲基蓝分光光度法; 悬浮物采用重量法; 盐度采用盐度计法; 溶解氧采用碘量法; 化学需氧量采用海碱性高锰酸钾法; 无机氮由亚硝酸盐、硝酸盐和氨盐组成, 其中亚硝酸盐采用萘乙二胺分光光度法, 硝酸盐采用锌-镉还原法, 氨盐采用次溴酸盐氧化法。
1.3 数据统计分析渔业资源密度(尾数、重量)估算方法: 设定拖网网具鱼类、甲壳动物和头足类尾数、重量逃逸率均为0.5。调查水域各测站拖网资源密度的估算采用扫海面积法, 渔业资源密度以各站拖网渔获量(重量、尾数)和拖网扫海面积来估算, 计算公式为
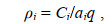 (1)
(1)式中, ρi 为第i站的资源密度(重量: kg/km2; 尾数: 103 ind./km2); Ci 为第i站的每小时拖网渔获量(重量: kg/h; 尾数: ind./h); ai为第i站的网具每小时扫海面积(km2/h) [网口水平扩张宽度(km)×拖曳距离(km)], 拖曳距离为拖网速度(km/h)和实际拖网时间(h)的乘积; q为网具捕获率=1
为方便统计与文字表达, 文中将尾数密度用丰度代替, 重量密度用生物量代替。
优势种在群落的数量和重量上占据显著比例。游泳动物优势种需综合其季节分布特点和个体大小差异, 在而相对重要性指数(Pinkas et al, 1971)能较好地反映游泳动物的优势种特征(朱鑫华等, 2002)。相对重要性指数计算公式如下:
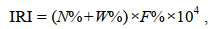 (2)
(2)式中, IRI为相对重要性指数; N%为某一物种尾数占总尾数的百分比; W%为该物种重量占总重量的百分比; F%为某一物种出现的站数占调查总站数的百分比(即出现频率)。以IRI > 500为优势种(刘金虎等, 2020)。
利用游泳动物群落香农威纳多样性指数(H’)、群落均匀度指数(J’)和丰富度指数(d)等来评价各个站位的生物多样性状况。
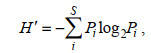 (3)
(3)式中, H′为物种多样性指数值; S为样品中的总种数; Pi为第i种的个体丰度(ni)与总丰度(N)的比值(ni/N)。
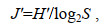 (4)
(4)式中, J′表示均匀度指数值; H′表示物种多样性指数值; S表示样品中总种数。
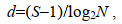 (5)
(5)式中, d表示丰富度指数值; S表示样品中的总种数; N表示群落中所有物种的总丰度。
采样站位分布图和群落相关参数的分布图采用软件ArcGIS Desktop 10.6绘制; 优势度计算使用了Microsoft Excel 2010; 采用多元统计软件PRIMER (Plymouth Routines in Multivariate Ecological Research) version 6.0计算群落中各个站位的Shannon-Wiener多样性指数、Pielou物种均匀度指数和Margalef物种丰富度指数; 采用PASW Statistics 18.0进行单因子ANOVA分析, 采用R包“psych”中的Pearson相关性分析和Canoco5.0软件的冗余分析(RDA)分析环境因子和群落参数相关性。
在相关性系数和多重双变量相关的P值的计算过程中使用了Bonferroni-Holm校正。首先对游泳动物丰度、生物量数据和环境因子数据进行对数[lg(x+1)]转换(宋晨等, 2022), 接着对游泳动物群落指数数据进行去趋势对应分析(Detrended Correspondence Analysis, DCA)得到轴梯度长度(lengths of gradient, LGA), 当LGA < 3时, 使用冗余分析(RDA); 当LGA < 4时, 使用典范对应分析(CCA); 当3 < LGA < 4时, 二者均可使用(马路阔等, 2022)。根据DCA结果, 本文适合使用线性模型的RDA分析游泳动物群落指数与环境因子的相关性。RDA排序图解读方法如下: 响应变量和解释变量箭头之间的夹角的余弦值即可反映出它们之间的相关性, 夹角越小, 相关性越大。响应变量之间、解释变量之间的关系亦如上。
2 结果 2.1 瓯江口游泳动物的物种多样性与物种数的空间分布在瓯江口的春、秋季的拖网调查中, 共采集到游泳动物78种, 隶属于5门8纲24目42科。其中, 鱼类15目19科36种, 占总采集种数的46.15%, 包括板鳃纲1种, 即光魟(Hemitrygon laevigata), 辐鳍鱼纲中以
瓯江口春季共获游泳动物48种, 其中脊索动物门鱼类(包括辐鳍纲和板鳃纲)共20种, 虾类10种, 蟹类9种, 头足类2种。秋季采获的游泳动物略多, 共66种, 其中鱼类32种, 虾类18种, 蟹类6种, 头足类3种。各站位间种类数差异均较大, 如在春季调查海域22号站位种类数最高(图 2)达27种, 包括鱼类9种, 而瓯江口附近的1号站位种类数最低仅出现9种, 鱼类仅有3种。秋季则以调查海域北侧, 乐清湾南部的4号站位种类数最高, 达28种, 包含鱼类14种, 瓯江口附近的1号站位种类数依旧最低, 仅8种, 包括鱼类4种(图 2)。
 |
| 图 2 瓯江口春、秋季游泳动物物种数量分布图 Fig. 2 The distribution of nekton species numbers in Oujiang River estuary in spring and autumn |
在春季底拖网调查渔获物丰度中, 鱼类比例最高, 为45.70%, 虾类次之, 达39.04%, 蟹类占13.32%, 头足类占0.41%, 其他占1.54%; 生物量组成中, 比例最高的类群为鱼类76.69%, 蟹类其次, 为9.32%, 虾类占5.21%, 头足类占5.04%, 其他占3.73%。秋季游泳动物丰度中, 鱼类所占比例最高, 为45.57%, 其次为虾类35.88%, 蟹类占16.45%, 头足类占1.18%, 其他占0.93%; 生物量组成中, 比例最高的类群为鱼类52.08%, 蟹类次之, 为22.94%, 较春季的比例有了大幅提高, 虾类占16.70%, 头足类占6.87%, 其他占1.42% (表 1)。
| 类群 | 春季 | 秋季 | |||||||
| 丰度/(×103 ind./km2) | 百分比/% | 生物量/(kg/km2) | 百分比/% | 丰度/(×103 ind./km2) | 百分比/% | 生物量/(kg/km2) | 百分比/% | ||
| 鱼类 | 2.82 | 45.7 | 85.46 | 76.69 | 4.66 | 45.57 | 60.91 | 52.08 | |
| 虾类 | 2.75 | 39.04 | 5.65 | 5.21 | 3.83 | 35.88 | 16.68 | 16.7 | |
| 蟹类 | 0.86 | 13.32 | 10.14 | 9.32 | 1.69 | 16.45 | 25.42 | 22.94 | |
| 头足类 | 0.04 | 0.41 | 5.13 | 5.04 | 0.1 | 1.18 | 6.79 | 6.87 | |
| 其他 | 0.1 | 1.54 | 3.74 | 3.73 | 0.08 | 0.93 | 1.55 | 1.42 | |
| 总计 | 6.56 | 100 | 110.13 | 100 | 10.36 | 100 | 111.34 | 100 | |
2021年春季渔获物的丰度和生物量均相对较低, 平均值分别为6.56×103 ind./km2 (3.23×103~16.27× 103 ind./km2)和110.13 kg/km2 (5.63~361.03 kg/km2)。其中, 鱼类均值为2.82×103 ind./km2和85.46 kg/km2, 虾类均值分别为2.75×103 ind./km2和5.65 kg/km2, 蟹类均值为0.86×103 ind./km2和10.14 kg/km2, 头足类均值为0.04×103 ind./km2和5.13 kg/km2, 其他均值为0.10×103 ind./km2和3.74 kg/km2 (表 1)。2020年秋季渔获物的丰度和生物量均略高于2021年春季, 平均值分别为10.36×103 ind./km2 (5.83×103~20.70×103 ind./km2)和111.34 kg/km2 (45.45~193.50 kg/km2)。其中, 鱼类均值为4.66×103 ind./km2和60.91 kg/km2, 虾类均值分别为3.83×103 ind./km2和16.68 kg/km2, 蟹类均值为1.69×103 ind./km2和25.42 kg/km2, 头足类均值为0.10×103 ind./km2和6.79 kg/km2, 其他均值为0.08× 103 ind./km2和1.55 kg/km2 (表 1)。
2021年春季, 丰度最大值出现在22号站位, 为16.27×103 ind./km2, 最小值出现在23号站位, 为3.23×103 ind./km2; 生物量最大值出现在8号站位, 为361.03 kg/km2, 最小值出现在1号站位, 为5.63 kg/km2 (表 2)。从平面分布上看, 调查海域丰度分布规律较均匀, 仅(图 3), 整体上丰度相对较低, 东南部22号站位出现大量的孔
| 站位 | 春季 | 秋季 | |||
| 丰度/(×103 ind./km2) | 生物量/(kg/km2) | 丰度/(×103 ind./km2) | 生物量/(kg/km2) | ||
| 1 | 3.68 | 5.63 | 12.34 | 154.67 | |
| 3 | 5.35 | 59.15 | 11.34 | 145.58 | |
| 4 | 11.11 | 189.51 | 14.86 | 193.50 | |
| 5 | 5.64 | 83.95 | 6.71 | 110.04 | |
| 8 | 3.33 | 361.03 | 6.58 | 80.70 | |
| 9 | 3.57 | 169.13 | 5.83 | 45.45 | |
| 14 | 6.19 | 23.67 | 7.92 | 71.04 | |
| 15 | 9.18 | 43.88 | 9.06 | 99.00 | |
| 19 | 5.03 | 38.90 | 10.73 | 66.73 | |
| 22 | 16.27 | 134.96 | 20.70 | 121.73 | |
| 23 | 3.23 | 145.85 | 5.84 | 97.45 | |
| 25 | 6.14 | 65.87 | 12.39 | 150.24 | |
| 平均 | 6.56 | 110.13 | 10.36 | 111.34 | |
 |
| 图 3 瓯江口春、秋季游泳动物丰度分布图 Fig. 3 The distribution of nekton density based on abundance in Oujiang River estuary in spring and autumn |
 |
| 图 4 瓯江口春、秋季游泳动物生物量分布图 Fig. 4 The distribution of nekton density based on biomass in Oujiang River estuary in spring and autumn |
2020年秋季, 丰度最大值出现在22号站位, 为20.70×103 ind./km2, 最小值出现在9号站位, 为5.83×103 ind./km2; 生物量最大值出现在4号站位, 为193.50 kg/km2, 最小值同样出现在9号站位, 为45.45 kg/km2 (表 2)。从平面分布上看, 调查海域丰度分布规律不明显(图 3), 东南部和北部海域丰度相对较高, 东南部海域主要是因为附近站位龙头鱼(Harpadon nehereus)出现得较多; 生物量方面, 北部靠近乐清湾的4号站和洞头列岛附近的22号站位生物量较高(图 4)。
2.3 瓯江口游泳动物优势种根据相对优势度指数(IRI), 2021年春季渔获物中鱼类优势种为孔
| 类群 | 种名 | 拉丁名 | 春季IRI | 秋季IRI |
| 鱼类 | 孔虎鱼 |
Trypauchen vagina | 1 862.58 | |
| 花鲈 | Lateolabrax japonicus | 1 698.41 | ||
| 棘头梅童鱼 | Collichthys lucidus | 1 144.45 | 622.30 | |
| 拉氏狼牙虎鱼 |
Odontamblyopus lacepedii | 688.29 | ||
| 刀鲚 | Coilia nasus | 617.10 | 732.59 | |
| 龙头鱼 | Harpadon nehereus | 934.87 | ||
| 甲壳动物 | 脊尾白虾 | Palaemon carinicauda | 1 658.33 | |
| 细巧仿对虾 | Batepenaeopsis tenella | 843.38 | ||
| 口虾蛄 | Oratosquilla oratoria | 2 763.12 | ||
| 三疣梭子蟹 | Portunus trituberculatus | 570.23 | 2 147.06 | |
| 日本蟳 | Charybdis japonica | 572.78 | 1 908.10 |
2020年秋季渔获物中鱼类优势种为龙头鱼、刀鲚和棘头梅童鱼, 虾类优势种有口虾蛄, 蟹类优势种有三疣梭子蟹和日本蟳(表 3)。口虾蛄为调查海域秋季最大优势种, 丰度为平均值为1.37×103 ind./km2, 最大值出现在4号站位, 为4.78×103 ind./km2, 1号站位未出现口虾蛄, 调查海域丰度分布规律不明显, 但北部乐清湾和南部丰度相对较高。生物量平均值为12.63 kg/km2, 最大值同样出现在4号站位, 为50.76 kg/km2; 调查海域生物量同样分布规律不明显, 在北部乐清湾和南部生物量相对较高。
2.4 瓯江口游泳动物群落指数的平面分布及其与环境因子相关性分析2021年春季丰度多样性指数(H')均值为2.97 (2.44~ 3.64), 均匀度指数(J')均值为0.78 (0.59~0.91), 丰富度指数(d)均值为1.11 (0.68~1.86) (表 4)。渔获物生物量多样性指数(H'-w)均值为2.58 (1.72~3.64), 均匀度指数(J'-w)均值为0.67 (0.42~0.85), 丰富度指数(d-w)均值为2.36 (1.18~3.67)。调查海域渔获物H'和H'-w各站位间差异不大, 分布较均匀。
| 站位 | 春季 | 秋季 | |||||||||||||
| 丰度 | 生物量 | 丰度 | 生物量 | ||||||||||||
| H' | J' | d | H'-w | J'-w | d-w | H' | J' | d | H'-w | J'-w | d-w | ||||
| 1 | 2.49 | 0.79 | 0.68 | 2.68 | 0.85 | 3.21 | 1.76 | 0.59 | 0.52 | 1.73 | 0.58 | 0.96 | |||
| 3 | 2.84 | 0.79 | 0.89 | 2.41 | 0.67 | 1.87 | 3.93 | 0.87 | 1.63 | 3.49 | 0.77 | 3.06 | |||
| 4 | 2.44 | 0.59 | 1.26 | 1.75 | 0.42 | 2.25 | 3.65 | 0.76 | 1.95 | 3.59 | 0.75 | 3.55 | |||
| 5 | 3.36 | 0.75 | 1.69 | 2.99 | 0.67 | 3.29 | 3.51 | 0.88 | 1.18 | 2.97 | 0.74 | 2.21 | |||
| 8 | 2.77 | 0.80 | 0.85 | 1.72 | 0.50 | 1.18 | 3.64 | 0.84 | 1.50 | 3.21 | 0.74 | 3.00 | |||
| 9 | 2.73 | 0.82 | 0.76 | 1.91 | 0.57 | 1.22 | 2.90 | 0.91 | 0.64 | 2.67 | 0.84 | 1.45 | |||
| 14 | 2.70 | 0.73 | 0.95 | 2.80 | 0.76 | 2.63 | 3.86 | 0.89 | 1.47 | 3.13 | 0.73 | 3.09 | |||
| 15 | 2.67 | 0.72 | 0.91 | 2.51 | 0.68 | 2.20 | 3.42 | 0.84 | 1.22 | 3.14 | 0.77 | 2.41 | |||
| 19 | 3.20 | 0.84 | 1.06 | 2.57 | 0.67 | 2.46 | 3.87 | 0.88 | 1.49 | 3.46 | 0.79 | 3.30 | |||
| 22 | 3.56 | 0.75 | 1.86 | 3.64 | 0.77 | 3.67 | 3.60 | 0.78 | 1.67 | 3.09 | 0.67 | 3.46 | |||
| 23 | 3.64 | 0.91 | 1.29 | 3.10 | 0.77 | 2.09 | 4.02 | 0.89 | 1.76 | 3.78 | 0.83 | 3.33 | |||
| 25 | 3.27 | 0.84 | 1.11 | 2.88 | 0.74 | 2.32 | 3.00 | 0.73 | 1.18 | 3.34 | 0.82 | 2.21 | |||
| 平均 | 2.97 | 0.78 | 1.11 | 2.58 | 0.67 | 2.36 | 3.43 | 0.82 | 1.35 | 3.13 | 0.75 | 2.67 | |||
2020年秋季H'均值为3.43 (1.76~4.02), J'均值为0.82 (0.59~0.91), d均值为1.35 (0.52~1.95) (表 4)。渔获物H'-w均值为3.13 (1.73~3.78), J'-w均值为0.75 (0.58~0.84), d-w均值为2.67 (0.96~3.55)。调查海域渔获物H'和H'-w除位于瓯江口里面的1号站位较低外, 其余站位分布较均匀。
将群落丰度、生物量以及基于丰度和生物量的群落香农威纳多样性指数、均匀度指数和丰富度指数与研究海域的环境因子, 包括有机碳、硫化物、活性磷酸盐、悬浮物、水深、盐度、溶解氧、化学需氧量、叶绿素a和无机氮含量进行Pearson相关性分析(图 5), 结果表明, 研究海域游泳动物群落多样性指数(H'和H'-w)与悬浮物的含量存在极显著的负相关关系, 随着悬浮物含量的上升, 这2个多样性指数值呈现显著的下降(P < 0.05), 其中悬浮物浓度与H'之间呈现极显著的负相关关系(Correlation=–0.75, P < 0.01)。除此之外, 游泳动物群落H'和H'-w与盐度存在较高的正相关关系, 但经过Bonferroni-Holm校正之后, 其相关性仅在0.1的水平上(0.05 < P > 0.1)。
 |
| 图 5 瓯江口游泳动物群落生态指数及环境因子相关性分析 Fig. 5 Correlation relationship between ecological indices of nekton assemblages and environmental factors in Oujiang River estuary 注: a. 相关性; b. 显著性水平 |
对生物的影响往往不是单个环境因素引起的, 而是多个环境因素综合产生的影响, 因此本文选择多元统计方法RDA来分析环境因子对瓯江口游泳动物群落指数的影响。通过前向选择(Forward selection)将环境因子排序, 利用Monte-Carlo置换检验(Permutations=499)评估环境变量对于解释群落指数变量的贡献, 选取对群落指数贡献高的环境因子绘制排序图, 分别为悬浮物、无机氮、水深、活性磷酸盐、叶绿素a和盐度(图 6)。RDA分析结果如下: 第一排序轴和所有排序轴存在显著差异(P=0.03 < 0.05), 说明RDA分析结果可信。RDA1特征值为0.750 9, 解释变量为40.6%, RDA2特征值为0.527 7, 解释变量为4.46%, 前两轴累计解释了45.06%的游泳动物群落指数与环境之间的关系。由图 6可知, 悬浮物是影响瓯江口游泳动物群落指数分布最大的环境因子, 其次为盐度和叶绿素a。其中, 游泳动物群落指数与悬浮物的含量存在显著的负相关关系, 与盐度和叶绿素a存在正相关关系, RDA分析结果与Pearson相关性分析结果相似, 影响游泳动物群落指数的主要环境因子为悬浮物和盐度。
 |
| 图 6 瓯江口游泳动物群落指数与环境因子RDA排序图 Fig. 6 RDA ordination graphs of community index and environmental factors in Oujiang River estuary 注: H': 丰度多样性指数; J': 丰度均匀度指数; d: 丰度丰富度指数; H'-w: 生物量多样性指数; J'-w: 生物量均匀度指数; d-w: 生物量丰富度指数 |
淡水径流是河口功能的强大驱动因素, 因而盐度梯度和其他理化因素的变化是河口的主要特征(Hobbie, 2000)。盐度可造成生境不稳定和群落扰动, 并与群落稳定性密切相关(Van Diggelen et al, 2016)。河口动物对底栖环境扰动反应灵敏, 是反映当地环境状况的良好指标(Qiu et al, 2018), 在河口生存的动物能够忍受盐度的大幅变化, 例如5~55之间的盐度变化也不会对河口动物造成致命影响(de Villiers et al, 1999)。因此, 随着盐度梯度的下降, 物种丰富度受到盐度梯度的抑制, 在8~10的盐度下物种多样性达到最低(Drake et al, 2002), 例如位于瓯江口的1号站物种数量在春秋季均为所有站位中最少, 其春季盐度仅12.2, 位于大门岛和霓屿岛, 中间的9号站的盐度次之, 其物种数亦是所有采样站位中, 仅多于瓯江口1号站的。物种数量最高的则为受洞头列岛庇护, 远离瓯江冲淡水影响的22号站(春季物种27种)以及乐清湾南部的4号站(秋季物种28种)。对照历史研究报道, 本研究在瓯江口海域共采集到游泳动物78种, 其中鱼类36种, 鱼类物种数远低于2007年夏秋的64种(徐兆礼, 2008)以及2018~2019年的84种(陈伟峰等, 2021), 游泳动物总物种数远高于2015年40种(刘志坚等, 2016), 但口足目2种和虾类16种与以往研究的口足目2种和虾类17种(徐兆礼等, 2009)相差无几。
历史研究中, 2015年采获的游泳动物数量最少, 这是由于该研究的采样站位主要设置于瓯江下游, 是针对淡水和咸淡水游泳动物的采集(刘志坚等, 2016); 瓯江口东侧尤其是洞头列岛以东海域, 受浙江沿岸水、黑潮次表层水和台湾暖流等海流水团的共同影响, 浮游生物饵料丰富, 是众多海洋游泳生物的产卵场和索饵场, 在此海域形成著名的温台渔场(李建生等, 2015)。采样区域越向东部外海扩展, 其能采集到的物种数量越多。5个站位位于洞头列岛以东且最东部采样站位达121.3°E的64种(徐兆礼, 2008), 而8个站位位于洞头列岛以西, 最东端达121.8°E以东即可采集到鱼类84种(陈伟峰等, 2021)。与附近海域相比, 采样区域广阔的研究能获得的物种数量也更高, 例如在温州湾春季的104种(李超男等, 2017)、乐清湾的128种(张琳琳等, 2019), 更是低于浙江中南部春季的166种游泳动物(蒋日进等, 2019)。
瓯江为源短流急的山溪性潮汐河流, 最大洪峰流量为22 800 m3/s, 最小流量仅为10.6 m3/s, 达洪枯流量比可达2 000倍, 平均输沙量为250万t, 最大浑浊带位于龙湾附近, 含沙量高达5~7 kg/m3 (王顺中等, 2006)。瓯江口附近海域的流场受到上游瓯江、各岩岛尤其瓯江口灵昆岛的分割引起的水流通道和域内众多大小沙洲的共同影响, 兼之霓屿岛与灵昆岛之间的灵霓北堤的阻隔, 瓯江径流分为南北两支, 北口径流沿着大门岛以南水道流向外海, 在瓯江口北侧大门岛以南海域存在一个巨大的冲淡水水团(徐兆礼, 2008); 而南口径流则向南流, 覆盖霓屿岛以南海域(姚炎明等, 1998)。两个南北冲淡水团基本覆盖了除乐清湾南部4号站以及洞头岛附近的22号站之外的大部分采样区域, 因此, 游泳动物的丰度在春季和秋季均以洞头岛附近的22号站为最大, 乐清湾南部的4号站次之。在生物量方面, 这两个站也处于较高水平, 但由于8号站采获到了个体较大的鮻和花鲈, 生物量更高, 而先前的研究结果也显示瓯江口洞头列岛外侧海域的游泳动物的密度较高(徐兆礼, 2008, 2009)。
3.2 瓯江口游泳动物优势种及其平面分布春季鱼类优势种为孔
春季虾类优势种有脊尾白虾和细巧仿对虾, 蟹类优势种有日本蟳和三疣梭子蟹, 秋季虾类优势种有口虾蛄, 蟹类优势种有三疣梭子蟹和日本蟳。口虾蛄(Oratosquilla oratoria)在每个季节的总渔获量中所占比例均远远高于其他各种类, 是整个东海口足类中的绝对优势种(卢占晖等, 2013)。在2021年春季的调查中, 口虾蛄平均丰度达24.43 ind./km2, 幼体所占比例高达81.8%, 且仅在4号和22号站位出现; 而秋季口虾蛄平均丰度达1.37×103 ind./km2, 幼体比例仅58.5%, 且分布于调查海域除瓯江口内的1号站位以外的所有站位, 口虾蛄也是瓯江口6月和9月的虾类第一优势种(徐兆礼等, 2009), 因此可以确定瓯江口附近海域是口虾蛄的育幼场和索饵场。本次调查与先前的研究一致, 日本蟳是瓯江口蟹类最重要的优势种, 三疣梭子蟹是最重要的经济品种(徐兆礼, 2009)。日本蟳在东海区广泛分布且可常年进行捕捞生产, 出现渔获量高峰是11月, 是该月份的第二大生物量优势种(俞存根等, 2005), 在2020~2021年的调查中, 日本蟳是11月份的第三大优势种, 其优势度仅次于口虾蛄和三疣梭子蟹。三疣梭子蟹在沿岸及河口水域产卵、索饵和育幼, 该物种在调查海域内分布范围最广, 出现的时间最长且对季节变化适应能力较强, 能够更好地利用环境资源, 在竞争中处于绝对优势(张琳琳等, 2019), 但在春季和冬季基础饵料生物不甚丰富的时候, 三疣梭子蟹会随着水温的降低其向深海区进行越冬洄游, 这就解释了三疣梭子蟹在春季(IRI=570.23)和秋季(IRI=2147.06) IRI值差异巨大的原因。
3.3 瓯江口游泳动物群落生态指数与环境因子的相关性根据Pearson相关性分析和RDA分析结果, 影响游泳动物群落指数的主要环境因子为悬浮物和盐度。在瓯江口研究海域, 悬浮物是影响游泳动物群落组成的最重要的环境因子。河口海域高浓度的悬浮物会降低水体中透明度和真光层厚度, 不利于浮游植物的光合作用, 进而会影响浮游动物及以其为饵料的生物的分布和生产力(李冠国等, 2004; 李开枝等, 2012), 直接决定着食物链顶端的游泳动物的数量分布(王娟娟等, 2016), 悬浮物与叶绿素a含量的相关关系分析的结果(Correlation=–0.407, P < 0.05)及RDA分析结果(二者夹角接近180°)也支持两者呈现显著的负相关关系。高浓度的悬浮物对游泳能力较弱的鱼类, 尤其是幼体影响很大(宋伦等, 2012), 是鱼类仔鱼丰度降低的最普遍因素之一。悬浮物的沉积会减少鱼类的饵料生物量, 降低鱼类的繁殖能力和鱼卵孵化率(周勇等, 2009), 并且影响了仔鱼的集群效应和抵御水流干扰的能力, 进而降低了游泳鱼类的种群密度(Wagener et al, 1985)。水体中悬浮泥沙可以影响鱼类心脏的每搏输出量和心输出量, 从而影响其正常的生理功能(Bunt et al, 2004; 宋伦等, 2012); 在悬浮物影响较大的海域, 悬浮物通过在鱼类等的鳃片、鳃丝及鳃小片等区域的沉积和聚集, 导致其窒息甚至死亡(Cordone et al, 1961; Wilber et al, 2001); 或者通过影响其进食行为而将游泳能力强的成鱼驱逐出该水域(高文斌等, 2009)。与此同时, 悬浮物中的颗粒有机物也是部分游泳动物的食物和能量来源(张铭汉, 2000); 另外, 高悬浮物的环境也会大幅降低被捕食者侵袭的危险概率, 从而使被庇护的游泳动物大量繁殖, 改变游泳动物的群落组成(Gradall et al, 1982), 因此, 悬浮物是影响游泳动物分布的重要因素(求锦津等, 2018; 张琳琳等, 2019), 高悬浮物的河口水域生物多样性(Blaber et al, 1997)和生产力(贾胜华等, 2017)要低于其他浑浊度相对较低的河口水域。
此外, 盐度与游泳动物的群落多样性也存在一定相关性。盐度可通过对狭盐性水生生物(如鱼类等游泳动物)的分布进行胁迫限制(Teske et al, 2003), 进而影响游泳动物群落的密度、丰富度和多样性(Gaston et al, 1998; Weslawski, 1999; Udalov et al, 2004)。在瓯江口海域, 海水悬浮物是影响并形成游泳动物群落的主要影响因子, 它与盐度等环境因子紧密联系, 共同影响和构建了河口近岸海域游泳动物物种数、密度和多样性较低, 而洞头列岛东侧、南侧和北侧等受瓯江冲淡水影响较小的水域游泳动物物种数、密度和多样性较高的地理分布格局。
4 结论(1) 瓯江口共采集到游泳动物78种, 其中鱼类是主要的游泳动物类群, 有36种, 甲壳动物28种, 头足类3种。
(2) 春季游泳动物优势种为孔
(3) 春季的渔获物的丰度远低于秋季, 而春季的生物量则与秋季的生物量相差不大。基于丰度和生物量的多样性指数(H')、均匀度指数(J')和丰富度指数(d)均为秋季高于春季。
(4) 在瓯江口, 水体悬浮物是影响游泳动物群落多样性和丰富度的最重要的环境因子, 它与盐度等环境因子综合作用, 共同形成了瓯江河口海域游泳动物群落的地理分布格局。
马路阔, 陈全震, 程方平, 等, 2022. 三门湾游泳动物群落结构与生物多样性研究. 海洋与湖沼, 53(2): 384-393 |
王顺中, 李浩麟, 2006. 瓯江口拦门沙航道回淤分析. 水利水运工程学报, (3): 18-24 |
王娟娟, 缴建华, 马丹, 等, 2016. 围填海吹填淤泥及悬浮物对天津海域海洋生物资源的急性毒性效应. 渔业科学进展, 37(2): 16-24 |
中国科学院中国动物志编辑委员会, 2008. 中国动物志: 硬骨鱼纲: 鲈形目(五): 虾虎鱼亚目. 北京: 科学出版社
|
中国海湾志编纂委员会, 1993. 中国海湾志: 第六分册: 浙江省南部海湾. 北京: 海洋出版社
|
卢占晖, 薛利建, 张亚洲, 2013. 东海口足类(Stomatopod)种类组成和数量分布. 自然资源学报, 28(12): 2159-2168 |
朱鑫华, 唐启升, 2016. 渤海鱼类群落优势种结构及其种间更替. 海洋科学集刊, (44): 159-168 |
刘志坚, 李德伟, 郭安托, 等, 2016. 瓯江口春夏季渔业生物种类组成及多样性. 浙江农业科学, 57(8): 1325-1327 |
刘金虎, 李毅, 曹亮, 等, 2020. 闽东近海游泳动物群落组成、资源量与生物多样性的时空格局研究. 海洋与湖沼, 51(4): 931-942 |
李开枝, 尹健强, 黄良民, 等, 2012. 珠江口伶仃洋海域底层游泳动物的季节变化. 生态科学, 31(1): 1-7 |
李建生, 严利平, 胡芬, 等, 2015. 温台渔场日本鲭的繁殖生物学特征. 中国水产科学, 22(1): 99-105 |
李冠国, 范振刚, 2004. 海洋生态学. 北京: 高等教育出版社
|
李超男, 水玉跃, 田阔, 等, 2017. 温州湾春季主要游泳动物生态位与种间联结性. 生态学报, 37(16): 5522-5530 |
求锦津, 王咏雪, 李铁军, 等, 2018. 舟山长白海域主要游泳动物生态位及其分化研究. 生态学报, 38(18): 6759-6767 |
宋伦, 杨国军, 王年斌, 等, 2012. 悬浮物对海洋生物生态的影响. 水产科学, 31(7): 444-448 |
宋晨, 孟周, 王晓波, 等, 2022. 2019-2020年夏季舟山海域浮游动物优势种生态位及其生态分化. 海洋学报, 44(10): 127-139 |
张铭汉, 2000. 胶州湾海水中悬浮体的分布及其季节变化. 海洋科学集刊, (42): 49-54 |
张琳琳, 蒋日进, 印瑞, 等, 2019. 乐清湾主要游泳动物空间生态位及其分化. 应用生态学报, 30(11): 3911-3920 |
陈伟峰, 叶深, 秦松, 等, 2021. 瓯江口海域鱼类群落结构及优势种RDA分析. 中国水产科学, 28(12): 1536-1547 |
林显鹏, 朱增军, 李鹏飞, 2010. 东海区龙头鱼摄食习性的研究. 海洋渔业, 32(3): 290-296 |
罗海舟, 张华东, 李鹏飞, 等, 2012. 东海区龙头鱼数量分布与渔业生物学现状分析. 浙江海洋学院学报(自然科学版), 31(3): 202-205, 233 |
周勇, 马绍赛, 曲克明, 等, 2009. 悬浮物对半滑舌鳎胚胎和初孵仔鱼的毒性效应. 渔业科学进展, 30(3): 32-37 |
俞存根, 宋海棠, 姚光展, 2005. 东海日本蟳的数量分布和生物学特性. 上海水产大学学报, 14(1): 40-45 |
姜亚洲, 林楠, 袁兴伟, 等, 2013. 象山港游泳动物群落结构及多样性. 生态学杂志, 32(4): 920-926 |
姜亚洲, 林楠, 袁兴伟, 等, 2015. 基于碳、氮稳定同位素技术研究象山港虾虎鱼类营养生态位. 生态学杂志, 34(6): 1579-1585 |
姜存楷, 1994. 温州市持续实行海蜇放流. 海洋信息, 9(S2): 42 |
姚炎明, 陈吉余, 陈永平, 等, 1998. 温州湾水流及其与地形的关系. 华东师范大学学报(自然科学版), (2): 61-68 |
贾胜华, 廖一波, 曾江宁, 等, 2017. 近岸典型生态系统大型底栖动物的次级生产力——以瓯江口、洞头列岛为例. 生态学报, 37(21): 7140-7151 |
徐兆礼, 2008. 夏秋季瓯江口海域鱼类数量的时空分布. 动物学报, 54(6): 981-987 |
徐兆礼, 2009. 瓯江口海域夏秋季蟹类数量分布与环境的关系. 水产学报, 33(2): 237-244 |
徐兆礼, 沈盎绿, 李新正, 2009. 瓯江口海域夏、秋季口足目和十足目虾类分布特征. 中国水产科学, 16(1): 104-112 |
高文斌, 刘修泽, 段有洋, 等, 2009. 围填海工程对辽宁省近海渔业资源的影响及对策. 大连水产学院学报, 24(S1): 163-166 |
蒋日进, 张琳琳, 徐开达, 等, 2019. 浙江中南部近岸海域游泳动物功能群特征与多样性. 生物多样性, 27(12): 1330-1338 |
樊伟, 周甦芳, 崔雪森, 等, 2003. 拖网捕捞对东海渔业资源种群结构的影响. 应用生态学报, 14(10): 1697-1700 |
BLABER S J M, FARMER M J, MILTON D A, et al, 1997. The ichthyoplankton of selected estuaries in Sarawak and Sabah: composition, distribution and habitat affinities. Estuarine, Coastal and Shelf Science, 45(2): 197-208 |
BUNT C M, COOKE S J, SCHREER J F, et al, 2004. Effects of incremental increases in silt load on the cardiovascular performance of riverine and lacustrine rock bass, Ambloplites rupestris. Environmental Pollution, 128(3): 437-444 |
CORDONE A J, KELLEY D W, 1961. The influences of inorganic sediment on the aquatic life of streams. California Fish and Game, 47(2): 189-228 |
DE VILLIERS C, HODGSON A, FORBES A, 1999. Studies on estuarine macroinvertebrates [M]//ALLANSON B R, BAIRD D. Estuaries of South Africa. Cambridge: Cambridge University Press: 167-208.
|
DRAKE P, ARIAS A M, BALDÓ F, et al, 2002. Spatial and temporal variation of the nekton and hyperbenthos from a temperate European estuary with regulated freshwater inflow. Estuaries, 25(3): 451-468 |
GASTON G R, RAKOCINSKI C F, BROWN S S, et al, 1998. Trophic function in estuaries: response of macrobenthos to natural and contaminant gradients. Marine and Freshwater Research, 49(8): 833-846 |
GRADALL K S, SWENSON W A, 1982. Responses of brook trout and creek chubs to turbidity. Transactions of the American Fisheries Society, 111(3): 392-395 |
HOBBIE J E, 2000. Estuarine Science: A Synthetic Approach to Research and Practice. Washington, DC: Island Press
|
PINKAS L, OLIPHANT M S, IVERSON I L K, 1971. Food habits of albacore, Bluefin tuna, and bonito in California waters. Department of Fish and Game Fish Bulletin, 152: 1-105 |
QIU B C, ZHONG X, LIU X S, 2018. Assessment of the benthic ecological status in the adjacent waters of Yangtze River Estuary using marine biotic indices. Marine Pollution Bulletin, 137: 104-112 |
TESKE P R, WOOLDRIDGE T H, 2003. What limits the distribution of subtidal macrobenthos in permanently open and temporarily open/closed South African estuaries? Salinity vs. sediment particle size. Estuarine, Coastal and Shelf Science, 57(1/2): 225-238 |
UDALOV A A, BURKOVSKII I V, MOKIEVSKII V O, et al, 2004. Changes in the general characteristics of micro-, meio-, and macrobenthos along the salinity gradient in the White Sea estuary. Oceanology, 44(4): 514-525 |
VAN DIGGELEN A D, MONTAGNA P A, 2016. Is salinity variability a benthic disturbance in estuaries?. Estuaries and Coasts, 39(4): 967-980 |
WAGENER S M, LAPERRIERE J D, 1985. Effects of placer mining on the invertebrate communities of interior Alaska streams. Freshwater Invertebrate Biology, 4(4): 208-214 |
WESLAWSKI J M, 1999. Influence of salinity and suspended matter on benthos of an Arctic tidal flat. ICES Journal of Marine Science, 56: 194-202 |
WILBER D H, CLARKE D G, 2001. Biological effects of suspended sediments: a review of suspended sediment impacts on fish and shellfish with relation to dredging activities in estuaries. North American Journal of Fisheries Management, 21(4): 855-875 |