 2023, Vol. 54
2023, Vol. 54

九龙江口肥水-缢蛏养殖塘厌氧氨氧化微生物活性研究

http://dx.doi.org/10.11693/hyhz20230100002
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 蔡露, 翁铂森, 詹力扬, 张介霞, 胡恭任, 于瑞莲, 闫钰. 2023.
- CAI Lu, WENG Bo-Sen, ZHAN Li-Yang, ZHANG Jie-Xia, HU Gong-Ren, YU Rui-Lian, YAN Yu. 2023.
- 九龙江口肥水-缢蛏养殖塘厌氧氨氧化微生物活性研究
- STUDY ON THE ACTIVITY OF ANAEROBIC AMMONIA OXIDIZING BACTERIA IN COMBINATION CULTURE SYSTEM OF FERTILIZER POOL AND SINONOVACULA CONSTRICTA CULTURE POND IN JIULONG RIVER ESTUARY
- 海洋与湖沼, 54(5): 1340-1350
- Oceanologia et Limnologia Sinica, 54(5): 1340-1350.
- http://dx.doi.org/10.11693/hyhz20230100002
文章历史
-
收稿日期:2023-01-04
收修改稿日期:2023-04-24
引用本文
蔡露, 翁铂森, 詹力扬, 张介霞, 胡恭任, 于瑞莲, 闫钰. 2023. 九龙江口肥水-缢蛏养殖塘厌氧氨氧化微生物活性研究[J]. 海洋与湖沼, 54(5): 1340-1350

CAI Lu, WENG Bo-Sen, ZHAN Li-Yang, ZHANG Jie-Xia, HU Gong-Ren, YU Rui-Lian, YAN Yu. 2023. STUDY ON THE ACTIVITY OF ANAEROBIC AMMONIA OXIDIZING BACTERIA IN COMBINATION CULTURE SYSTEM OF FERTILIZER POOL AND SINONOVACULA CONSTRICTA CULTURE POND IN JIULONG RIVER ESTUARY[J]. Oceanologia et Limnologia Sinica, 54(5): 1340-1350.
九龙江口肥水-缢蛏养殖塘厌氧氨氧化微生物活性研究
1. 华侨大学化工学院 福建厦门 361000;
2. 自然资源部第三海洋研究所海洋大气化学与全球变化重点实验室 福建厦门 361000
2. 自然资源部第三海洋研究所海洋大气化学与全球变化重点实验室 福建厦门 361000
摘要:缢蛏串联养殖经由饵料藻类养殖塘(俗称肥水塘)投放适量含氮化合物以提高生产效率, 了解此过程中的沉积物氮转化对调控和管理河口养殖塘氮的收支具有重要意义。以福建九龙江口肥水-缢蛏养殖塘为研究对象, 运用同位素示踪技术和高通量测序技术探究缢蛏塘和肥水塘沉积物厌氧氨氧化速率及微生物群落结构, 并分析其环境调控机制。结果表明, 肥水塘沉积物厌氧氨氧化速率为(1.76±0.11) nmol/(g·h), 相对贡献为9.16%±0.40%, 显著高于缢蛏塘(P < 0.05)。缢蛏塘和肥水塘厌氧氨氧化微生物主要由Candidatus Scalindua、Ca. Kuenenia、Ca. Brocadia和Ca. Anammoximicrobium属组成。其中, Ca. Kuenenia属为养殖塘的优势菌属。沉积物厌氧氨氧化微生物多样性和丰富度呈显著的空间分布特征, 肥水塘中生物多样性更高(P < 0.05)。厌氧氨氧化微生物活性和群落结构差异主要受养殖土地利用方式、溶解氧(DO)、NH4+和NO3-含量等环境因子的调控。该研究在一定程度上揭示了养殖环境中厌氧氨氧化过程的重要性, 填补了河口养殖湿地氮回归大气的认知空白。
关键词:厌氧氨氧化 养殖塘 速率 群落结构 影响因素
STUDY ON THE ACTIVITY OF ANAEROBIC AMMONIA OXIDIZING BACTERIA IN COMBINATION CULTURE SYSTEM OF FERTILIZER POOL AND SINONOVACULA CONSTRICTA CULTURE POND IN JIULONG RIVER ESTUARY
1. School of Chemical Engineering, Huaqiao University, Xiamen 361000, China;
2. Key Laboratory of Global Change and Marine-Atmospheric Chemistry, Third Institute of Oceanography, MNR, Xiamen 361000, China
2. Key Laboratory of Global Change and Marine-Atmospheric Chemistry, Third Institute of Oceanography, MNR, Xiamen 361000, China
Abstract: A tandem culture system was designed to produce razor clam Sinonovacula constricta in pond with an upstream culture pond that was used to cultivate algae as bait for the razor clam in Jiulong River estuarine area, Fujian, SE China. A certain amount of nitrogen compounds was put into the algae pond for nurturing alga. The nitrogen (N) budget in water and sediment of the tandem system was studied to for best practice. The rates and microbial community structure of anaerobic ammoxidation (anammox), and their response to the ambience factor were studied using 15N isotope-tracing technology and high-throughput sequencing methods. Results show that the potential rate of anammox in sediment of the algae ponds was (1.76±0.11) nmol/(g·h), and the relative contribution rate to anaerobic ammoxidation was 9.16%±0.40%, which was significantly higher than of the S. constricta pond. Anammox bacteria in the S. constricta pond and the algae pond were dominated by Ca. Scalindua, Ca. Kuenenia, Ca. Brocadia, and Ca. Anammoximicrobium. The diversity and richness of anammox bacteria in sediment of the algae pond were significantly higher than those of the S. constricta pond (P < 0.05). Environmental factors including DO, NH4+, and NO3- contents controlled the anammox microbial community structure of the system. This study revealed the N utilization and removal processes of anammox, and filled the gap in the knowledge of N return into the atmosphere in estuarine aquaculture wetland.
Key words:
anammox aquaculture ponds nitrogen removal rates community structure impact factor
氮素是自然生态系统中重要的营养元素, 其生物地球化学循环是保证陆地和海洋环境安全的重要环节(祝贵兵, 2020)。随着滨海养殖业迅猛发展, 过量施用含氮饲料影响了沉积物氮循环过程, 造成严重的生态环境问题, 如水体富营养化、生物多样性丧失等(Wu et al, 1994)。反硝化和厌氧氨氧化作用是脱氮的两个重要过程(Mulder et al, 1995), 但反硝化过程会产生中间产物氧化亚氮(N2O), 其对温室效应的影响是二氧化碳(CO2)的298倍(Li et al, 2021), 尤其是在高氮负荷河口(Wells et al, 2018)。厌氧氨氧化在脱氮过程中减少了NO、N2O等温室气体的产生, 是一种更为经济有效且环保的途径(Suenaga et al, 2021)。因此, 探究滨海河口养殖塘沉积物厌氧氨氧化活性对提高水产养殖污染物控制和管理效率以及合理利用滨海湿地资源具有重要的理论意义。
目前, 共发现6个厌氧氨氧化微生物属(Schmid et al, 2005; Bonaglia et al, 2017), 包括Candidatus Brocadia、Ca. Scalindua、Ca. Kuenenia、Ca. Jettenia、Ca. Anammoximicrobium moscowii和Ca. Anammoxoglobus。虽然它们在生理、代谢和结构特征方面十分相似, 但其生存环境和生态位差异显著, 具有高度的栖息地特异性(Dale et al, 2009)。例如, 在海洋生态系统中Ca. Scalindua的丰度更高, 在高盐度环境中占绝对优势(Awata et al, 2013)。有研究报道, 海洋沉积物中厌氧氨氧化活性较强, 贡献率占40%~60% (Thamdrup et al, 2002; Brandsma et al, 2011), 而淡水湖泊、河口和自然土壤中以Ca. Brocadia、Ca. Kuenenia为主(Humbert et al, 2010), 其厌氧氨氧化相对贡献仅10%~20% (Shen et al, 2017)。Ca. Jettenia则较多存在于生物反应器和污水处理厂中(Quan et al, 2008; Sonthiphand et al, 2014)。当前, 对水产养殖塘也开展了一些研究, 养殖生态系统不同于自然环境, 受人为管理干扰较大, 氮负荷较高(杨雪琴等, 2018)。Shen等(2016)对江苏某淡水养殖塘沉积物厌氧氨氧化的研究中发现其速率为3.70~19.40 nmol/(g·d), 相对贡献为1.20%~15.30%, 并且Ca. Brocadia属为其优势属。另外, 对菲律宾蛤仔海水养殖塘有贝区和无贝区沉积物研究发现有贝区厌氧氨氧化速率为0.000 92 μmol/(kg·h), 且埋栖型贝类的扰动会影响沉积物厌氧氨氧化反应(朱文君等, 2022)。因此, 受微生物对环境基质的反应不同, 厌氧氨氧化作用及其相对贡献具有很强的不确定性(Teixeira et al, 2016)。滤食性贝类滩涂养殖具有明显的生态学效应, 其栖息在咸淡水交汇的河口附近和沿海砂泥底部, 能够通过调整其生理活动以适应环境的改变(吕昊泽等, 2016), 并影响系统的物质循环和能量流动, 进而影响微生物活性(柴欣如等, 2022)。目前, 对肥水-缢蛏养殖塘这一特殊养殖模式下沉积物厌氧氨氧化过程的研究还鲜见报道, 近海养殖区沉积物中厌氧氨氧化微生物的活性及分布需要更系统和深入地分析。
基于以上, 本文选择福建近海九龙江口肥水-缢蛏养殖塘为研究对象, 采用15N同位素示踪技术和高通量测序技术揭示肥水-缢蛏养殖模式下沉积物中厌氧氨氧化速率及微生物群落结构, 并应用统计分析方法探讨驱动厌氧氨氧化活性和微生物群落变化的主要环境因子, 对滨海养殖环境下的脱氮过程进行初步研究, 为保护近海水域生态环境、缓解全球气候变暖提供科学依据。
1 材料与方法 1.1 研究区概况九龙江口是福建典型的入海河口之一, 位于我国暖热湿润的亚热带东南沿海地区, 年均气温19.6~21.0 ℃, 年均降水量1 400~2 000 mm (程金等, 2023), 降水多集中在3~9月, 是红树林的主要分布地区之一(Yang et al, 2018)。红树林能提供优质的自然饵料和良好的庇护场所, 为该沿海地区水产养殖提供优越的条件(Alongi et al, 2005; 王文卿等, 2021)。缢蛏养殖由两个相邻的池塘组成, 即肥水塘和缢蛏塘, 肥水塘体积是缢蛏塘的3~5倍。较大的肥水塘主要用于施加肥料(碳酸氢铵肥料、有机无机复混肥等)以培养藻类作为饵料供缢蛏食用。缢蛏塘主要为蛏养殖场所, 接受肥水塘的肥水以及潮水中有机颗粒物。两塘之间设置闸门以便向缢蛏塘注入肥水。养殖周期通常是10月投放蛏苗, 次年5月全部收获, 5~7月缢蛏塘会进行底泥整塘清淤。养殖期间, 每天凌晨将缢蛏塘水排干, 早上再打开阀门使肥水塘的肥水流入缢蛏塘, 流入量仅占整个肥水塘池水的1/3或更低。肥水塘中水位下降至一定程度后打开阀门使对岸九龙江水流入灌满肥水塘, 并投放肥料, 投放量根据缢蛏的生长及摄食情况调整。
1.2 样品采集和处理于2021年4月选择了九龙江口紫泥红树林区周边的缢蛏-肥水养殖塘(24°27′28″N, 117°54′45″E; 图 1), 分别采集了5个缢蛏塘(C1、C2、C3、C4、C5)和4个肥水塘(Z1、Z2、Z3、Z4)的沉积物(0~10 cm)和上覆水样品共9个。采样时各池塘采集2 L水样于聚乙烯塑料瓶中密封, 并分别于每个塘中央及四周随机选取表层沉积物充分混匀作为一个塘的沉积物样品。混匀后立即置于聚乙烯密封袋中, 排尽空气后密封, 用冰盒储存带回实验室。一部分样品置于4 ℃冰箱保存, 并尽快进行理化参数的测定和同位素实验, 另一部分置于−80 ℃冰箱保存, 用于提取DNA进行分子生物学分析。
 |
| 图 1 养殖塘采样点分布 Fig. 1 Distribution of sampling site |
选择沉积物上覆水溶解氧(DO)、盐度(S)和沉积物pH值、氨氮(NH4+)、硝氮(NO3-)、亚硝氮(NO2-)、总氮(TN)、总碳(TC)和总硫(TS)为基本理化指标进行检测。沉积物上覆水DO和S在采样现场使用DZB-718L型便携式多参数分析仪测定。沉积物样品采用《土壤农化分析》中的方法进行相应测定(鲍士旦, 2000), 沉积物pH值采用酸度计测定(土水比1︰2.5)。NH4+、NO3-和NO2-的测定为采用2 mol/L的KCl溶液浸提沉积物样品1 h, 将浸提液用0.45 μm膜过滤后用连续流动分析仪(AAA3, Seal Analytical GmbH, 德国)测定, 沉积物TN、TC和TS利用元素分析仪(Elementar, Vario Micro, 德国)进行测定。沉积物总有机碳(TOC)采用总有机碳分析仪(TOC-LCPH, 日本岛津)测定。所有理化指标都选用3组平行样品进行测定, 结果取3组平行样品的平均值。
1.4 厌氧氨氧化速率和反硝化速率测定采用15N同位素示踪来测定反硝化和厌氧氨氧化反应速率及对N2产生的贡献。将50 g沉积物样品装入1 L的气袋中, 并加入250 mL经氦气曝气后的原位水, 混合制成均匀泥浆, 封口并将袋中空气通过气孔排尽。恒温摇动培养3天左右至氧气和NO3-耗尽。按实验分组在厌氧操作箱中分别加入5 mL未标记或15N标记溶液(15N终浓度为100 μmol/L): ①阴性对照, 只加(15NH4)2SO4; ②阳性对照, (15NH4)2SO4和Na14NO3; ③实验组: 添加Na15NO3。放入恒温培养箱中反应并按0、3、6、9、12 h时间点于厌氧操作箱内取样至12 mL真空玻璃管中, 并注入0.2 mL 7 mol/L氯化锌溶液终止反应。所加试剂均在配制时通过样品瓶充气抽真空清洗系统进行除氧。处理完的样品避光倒置保存, 于膜进样质谱仪(membrane inlet mass spectrometry, MIMS)上测定水中溶解的29N2和30N2的浓度, 29N2和30N2浓度的计算公式如下(谢成军等, 2020):
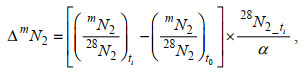 (1)
(1)式中, ∆mN2表示29N2和30N2的产生浓度; 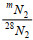
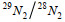
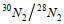
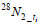
29N2和30N2产生速率通过各自氮气产生量对时间的回归得到, 反硝化和厌氧氨氧化速率以及相对贡献计算过程以处理③为例说明(Thamdrup et al, 2002; 赵永强等, 2014):
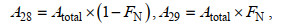 (2)
(2)式中, A28和A29分别表示厌氧氨氧化过程中各自N2的产生量; FN为所购药品K15NO3(阿拉丁, 上海)中15N的比例(FN=0.995)。处理③中反硝化反应产生的N2应包括28N2、29N2和30N2:
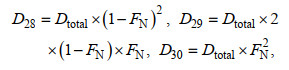 (3)
(3)式中, D28、D29和D30分别表示反硝化中各N2产生量。用P29和P30分别表示MIMS测得的各自N2总量, 结合公式(4)和(5), 且D30=P30, 从而可以得到:
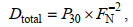 (4)
(4)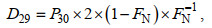 (5)
(5)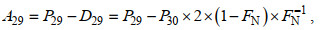 (6)
(6)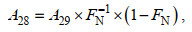 (7)
(7)叠加得
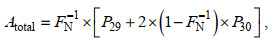 (8)
(8)Dtotal和Atotal分别为各样品中反硝化和厌氧氨氧化反应的N2产生量, 并对采集时间点进行线性回归, 即可得到该样品的反硝化和厌氧氨氧化速率。
反硝化和厌氧氨氧化各自的脱氮贡献率(Rd和Ra)可由下列公式计算得出:
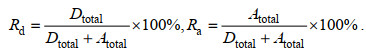 (9)
(9)称取0.33 g冷冻干燥的沉积物样品, 使用FastDNA SPIN Kit for Soil土壤DNA提取试剂盒提取沉积物样品中的总DNA, 并用Nano Drop 2000 UV-Vis Spectrophotometer (Thermo Fisher Scientific, USA)测定DNA浓度。
厌氧氨氧化微生物特异性16S rRNA基因(AMX)采用巢式PCR扩增的方法(Wang et al, 2012; 王衫允等, 2012), 以提取的DNA为模板, 先利用引物pla46F(5'-GGATTAGGCATGCAAGTC-3')和630R(5'-CAKAAAGGAGGTGATCC-3')对浮霉菌门进行PCR扩增, 第二步扩增则以第一步PCR产物为模板, 使用厌氧氨氧化微生物特异性引物Amx368F(5'-TTC GCAATGCCCGAAAGG-3')和Amx820R(5'-AAAA CCCCTCTACTTAGTGC CC-3')进行厌氧氨氧化微生物特异性16S rRNA基因的PCR扩增。PCR反应体系为30 μL, 其中, 2×Robust PCR Master Mix 15 μL, 10 mmol/L正反引物各1 μL, DNA模板2 μL, 用无核酸酶纯水补足至30 μL。扩增所得产物通过Hiseq 2500平台(Illumina, SanDiego, CA, USA)进行测序(派森诺生物有限公司, 上海)。高通量测序结果提交至GenBank(登录号为PRJNA914860)。
1.6 数据处理与分析高通量测序得到的原始序列经质控后, 利用Mothur软件(https://www.mothur.org/)去除低质量序列、及嵌合体, 过滤后得到高质量序列; 在97%相似度水平对高质量序列聚类, 并分别输出代表序列和操作分类单元(operational taxonomic unit, OTU)表。利用RDP classifier (http://rdp.cme.msu.edu/)对序列进行物种分类注释, 使用QIIME程序计算厌氧氨氧化微生物特异性16S rRNA基因的多样性指数。
使用SPSS 20.0 (SPSS Inc., Chicago, Ⅲ, USA)分析检验不同沉积物样品环境因子及反硝化和厌氧氨氧化速率显著性差异(P < 0.05)。使用Canoco (V5.0)软件进行去趋势对应分析(DCA), 根据其结果判断采用冗余分析(redundancy analysis, RDA)以评估环境因子对反硝化和厌氧氨氧化速率的影响。基于Bray-Curtis差异度指数采用主坐标分析(Principle Co-ordinate Analysis, PCoA)比较不同样品厌氧氨氧化微生物群落结构差异; 利用R语言(V4.1.3)进行Mantel tests相关性分析探究缢蛏塘和肥水塘厌氧氨氧化微生物群落结构与环境因子之间的相关性; 通过删除不显著路径, 并比较AIC值, 筛选出影响厌氧氨氧化速率和微生物群落的主要环境因子, 使用R语言piecewiseSEM包构建结构方程模型(structural equation model; SEM)(Huang et al, 2020; 石亚飞等, 2022)。利用Origin 2021软件绘图。
2 结果 2.1 养殖塘理化性质养殖塘理化性质如表 1所示。缢蛏塘和肥水塘上覆水DO水平在6.10~11.53 mg/L之间, 肥水塘DO水平显著高于缢蛏塘(P < 0.05)。缢蛏塘盐度(26.80±0.26)显著高于肥水塘(22.88±0.33)。所有沉积物样品pH值范围为7.69~8.30, 偏碱性。沉积物中氮形态以NH4+为主, 含量为38.19~51.07 mg/kg, 且NH4+含量在缢蛏塘和肥水塘中差异显著, 肥水塘中NH4+和NO3-含量平均值分别为(47.6±3.33)和(7.65±1.09) mg/kg, 均高于缢蛏塘(39.78±1.46)和(5.06±0.98) mg/kg (P < 0.05)。各采样点NO2-、TN和TS含量无明显差异, 范围分别为0.81~1.21 mg/kg、1.1~2.2 g/kg和7.59~16.22 g/kg。缢蛏塘和肥水塘碳氮比无显著差异, 但肥水塘中TC含量[(19.35±3.10) g/kg]和TOC含量[(14.90±0.45) g/kg]均显著高于缢蛏塘[TC: (14.08±1.48) g/kg; TOC: (9.60±0.73) g/kg; P < 0.05], 表明沉积物环境因子对养殖土地利用变化的响应十分敏感。
表 1 采样点沉积物物理化学参数 Tab. 1 Physicochemical parameters of the sampling site
| 样品 | 上覆水 | pH | 积物 | ||||||||
| DO/(mg/L) | S | NH4+/(mg/kg) | NO3-/(mg/kg) | NO2-/(mg/kg) | TN/(g/kg) | TOC/(g/kg) | TC/(g/kg) | TS/(g/kg) | C: N | ||
| C1 | 6.10 | 26.40 | 8.02 | 39.31 | 3.82 | 0.81 | 2.20 | 10.20 | 19.90 | 11.66 | 9.05 |
| C2 | 6.30 | 26.70 | 8.21 | 42.49 | 5.85 | 1.04 | 1.10 | 9.80 | 11.60 | 7.59 | 10.55 |
| C3 | 6.70 | 27.20 | 7.69 | 39.05 | 5.96 | 0.98 | 1.10 | 10.50 | 14.10 | 13.22 | 12.82 |
| C4 | 6.50 | 26.90 | 8.14 | 38.19 | 3.41 | 0.86 | 1.70 | 8.90 | 11.30 | 11.75 | 6.65 |
| C5 | 6.80 | 26.80 | 8.30 | 39.88 | 6.27 | 1.21 | 1.70 | 8.60 | 13.50 | 14.51 | 7.94 |
| Z1 | 11.06 | 23.10 | 8.27 | 47.82 | 7.69 | 1.16 | 2.20 | 15.10 | 19.90 | 11.66 | 9.05 |
| Z2 | 10.42 | 23.00 | 8.25 | 49.33 | 7.86 | 1.21 | 1.90 | 14.30 | 16.80 | 16.22 | 8.84 |
| Z3 | 11.53 | 23.10 | 8.33 | 51.07 | 9.13 | 1.18 | 2.20 | 14.70 | 20.40 | 15.63 | 9.27 |
| Z4 | 10.54 | 22.30 | 8.04 | 42.19 | 5.93 | 0.81 | 1.90 | 15.50 | 20.30 | 10.44 | 10.68 |
| 注: DO: 溶解氧; S: 盐度; NH4+: 氨氮; NO3-: 硝氮; NO2-: 亚硝氮; TN: 总氮; TC: 总碳; TS: 总硫; C: N表示TC与TN的比值。 | |||||||||||
缢蛏塘和肥水塘沉积物厌氧氨氧化和反硝化速率及厌氧氨氧化相对贡献如图 2所示, 缢蛏塘和肥水塘沉积物厌氧氨氧化速率范围分别为0.94~1.26和1.61~1.91 nmol/(g·h), 分别占总氮气生成量的6.09%~ 8.06%和8.73%~9.63%。肥水塘厌氧氨氧化速率[(1.76±0.20) nmol/(g·h)]显著高于缢蛏塘[(1.07±0.11) nmol/(g·h)]。两塘沉积物反硝化速率范围为12.06~ 18.26 nmol/(g·h)。缢蛏塘和肥水塘沉积物厌氧氨氧化速率和相对贡献表现出显著的养殖土地利用差异(P < 0.05)。RDA分析表明(图 3), DO、S、pH、NH4+、TC和NO3-含量是影响缢蛏塘和肥水塘厌氧氨氧化速率的主要因素。
 |
| 图 2 缢蛏塘和肥水塘沉积物厌氧氨氧化速率和相对贡献(a)及反硝化速率(b) Fig. 2 Potential anammox rates and relative contribution (a), and the denitrification rates (b) of sediments in the Sinonovacula constricta ponds and the breeding ponds |
 |
| 图 3 沉积物厌氧氨氧化速率与环境因子的冗余分析 Fig. 3 Redundancy analysis of anammox rate and environmental factors in sediments 注: DNF: 反硝化速率; ANA: 厌氧氨氧化速率; Ra: 厌氧氨氧化相对贡献率 |
沉积物厌氧氨氧化微生物的所有序列进行聚类共得到2 945个OTUs。对缢蛏塘和肥水塘沉积物样品进行α多样性指数统计分析, 通过Chao1、Shannon和Simpson指数评估厌氧氨氧化微生物群落多样性和丰富度, 结果见图 4。两塘沉积物厌氧氨氧化微生物多样性差异显著(P < 0.05)。并在OTU水平上计算了缢蛏塘和肥水塘沉积物样品的Bray-Curits差异度矩阵, 进行厌氧氨氧化微生物的群落排序, PCoA分析如图 5a所示, 可以发现缢蛏塘和肥水塘的样品距离较远, 表明其厌氧氨氧化微生物的群落结构差异较大。根据每个OTU的代表序列在系统发育树中的位置, 对代表序列对应的OTU的样品序列进行物种注释并计算每个样品中属水平上厌氧氨氧化微生物物种组成的相对丰度(图 5b), 结果显示其序列隶属于四个属, 分别为Ca. Scalindua、Ca. Kuenenia、Ca. Brocadia和Ca. Anammoximicrobium。不同样点的厌氧氨氧化微生物的物种组成差异不大, Ca. Kuenenia在所有样品中所占比例最大, 为优势菌属。
 |
| 图 4 缢蛏塘(CT)和肥水塘(ZT)沉积物样品厌氧氨氧化微生物AMX基因多样性指数 Fig. 4 Diversity index of anammox 16S rRNA gene sequence of sediments in the Sinonovacula constricta ponds and the breeding ponds 注: 不同小写字母标识表示缢蛏塘和肥水塘间差异显著(P < 0.05) |
 |
| 图 5 缢蛏塘和肥水塘沉积物厌氧氨氧化微生物群落PCoA分析图(a)和物种组成图(b) Fig. 5 Principal Co-ordinates Analysis and species composition of anammox microbial communities in sediments of the Sinonovacula constricta pond and the breeding ponds |
选取厌氧氨氧化微生物属水平组成相对丰度矩阵、多样性指数矩阵及前120个OTU数据矩阵与环境因子进行Mantel tests相关性分析, 以探究环境因子对沉积物厌氧氨氧化微生物群落的影响。由图 6可知, 厌氧氨氧化微生物物种组成分布与NH4+ (R=0.35, P=0.04)相关性显著。厌氧氨氧化微生物多样性与DO (R=0.24, P=0.04)和NO3- (R=0.45, P=0.03)显著相关, 与NH4+(R=0.53, P=0.002)呈极显著相关。通过分析筛选得到影响厌氧氨氧化速率和微生物群落的主要环境因子构建结构方程(SEM)模型, 如图 7所示, 养殖土地利用方式对DO和NH4+浓度具有极显著影响, 同时NH4+含量是厌氧氨氧化微生物多样性的主要影响因子。
 |
| 图 6 沉积物厌氧氨氧化微生物与环境因子Mantel tests相关性分析 Fig. 6 The mantel tests correlation analysis between community of anammox bacteria and chemical characters of sediments 注: 相关性大小与线的粗细呈正比; 线的颜色表示显著性水平的P值大小 |
 |
| 图 7 使用piecewiseSEM包进行环境因子对沉积物厌氧氨氧化微生物群落结构影响的结构方程模型分析 Fig. 7 The structural equation model (SEM) of the effects of environmental factors on anammox microbial community structure based on the piecewiseSEM package 注: 左图中红色、蓝色分别表示显著正、负的路径关系, 数值为标准化路径系数, 箭头粗细表示标准化路径系数的相对大小。*: P < 0.05; **: P < 0.01; ***: P < 0.001 |
厌氧氨氧化微生物广泛分布于自然生态系统中, 其介导的脱氮过程能够有效减少NO和N2O等温室气体的产生(Suenaga et al, 2021), 对控制全球气候变暖发挥着关键性作用。本研究区域沉积物厌氧氨氧化速率范围为0.94~1.91 nmol/(g·h), 相对贡献范围为6.09%~9.63%, 与农田土壤(Zhu et al, 2011)、潮白河沉积物(彭强等, 2021)的活性范围相近, 但显著低于九龙江口红树林湿地沉积物脱氮贡献(Cao et al, 2017)。另外, 经高通量测序分析发现缢蛏塘沉积物厌氧氨氧化微生物多样性不高, 进一步证实了功能性较强的环境中微生物多样性偏低(Loreau et al, 2001; 刘璐, 2016)。另外, 适应于淡水环境中的Ca. Kuenenia属在本研究中相对丰度占比最高, 表明了研究区域滨海河口养殖塘的特殊性, 连接着陆地和海洋(洪义国等, 2019; Ya et al, 2021)。养殖饲料中含有甲酸和丙酸, 作为抑制性饲料添加剂防止饲料腐败, 其也是养殖动物肠道中蛋白质及碳水化合物发酵的主要产物, Ca. Anammoximicrobium属存在于养殖塘中, 能够氧化甲酸和丙酸(Kartal et al, 2007; 张亦弛等, 2022)。
养殖土地利用方式、DO、NH4+和NO3-等被认为是影响本研究沉积物厌氧氨氧化微生物的主要环境因素。蓄水养藻的肥水塘上覆水盐度显著低于缢蛏塘, 推测可能由于周期性潮水盐度变化和采样时短期的降雨导致。缢蛏捕食以及扰动行为增加了水体浊度, 降低了水体光合作用(丁维新等, 2020), DO水平显著低于肥水塘。另外, 肥水塘中藻类的光合作用以及缢蛏塘中缢蛏的扰动作用会促使两塘中含氧的上覆水进入沉积物, 使得厌氧氨氧化在微厌氧的条件下发生(Cheng et al, 2014), 与Zhu等(2011)在水稻田的研究中发现DO的可利用性是厌氧氨氧化发生的重要因素的结论一致。肥水塘沉积物厌氧氨氧化速率[1.61~1.91 nmol/(g·h)]和脱氮相对贡献(8.73%~9.63%)显著高于缢蛏塘。并且, 肥水塘沉积物中厌氧氨氧化微生物多样性丰富, 群落结构复杂。有研究表明, 养殖动物扰动行为会改变沉积物性质和结构, 从而影响微生物群落(Kim et al, 2017)。厌氧氨氧化微生物对环境要求较高(Dalsgaard et al, 2005), 因此, 每日频繁交换水、养殖动物扰动和养殖周期底泥翻新等活动可能使得缢蛏塘沉积物厌氧氨氧化微生物活性受抑制, 从而脱氮速率显著低于营养盐充足、微生物多样性复杂且沉积环境相对稳定的肥水塘。NH4+浓度与厌氧氨氧化微生物群落呈显著正相关。底物中NH4+的可利用性是影响厌氧氨氧化活性的关键控制因素(Risgaard-Petersen et al, 2004; Shen et al, 2014), 可为厌氧氨氧化反应的发生提供底物。Zhu等(2011)对稻田土壤的研究中也证实较高的NH4+含量会促进厌氧氨氧化微生物活性。功能微生物与相应功能速率间的相关性并不显著可能是由于河口养殖受人为影响大、环境条件波动范围大, 限制了微生物生长活性和生物活动(Zheng et al, 2016)。
值得注意的是, 目前已有大量研究表明厌氧氨氧化具有较强的时空异质性(Shan et al, 2016; 毛铁墙等, 2020; Wu et al, 2021)。Zhang等(2016)研究指出, 阳澄湖淡水养殖塘养殖期沉积物厌氧氨氧化脱氮贡献显著低于非养殖期, 并且厌氧氨氧化微生物丰度随养殖季节发生显著变化。受养殖温度、养殖时期、季节和养殖类型等因素的影响, 氮转化速率及其相关功能微生物和功能基因的数量及群落结构对其的响应规律会受到取样时间的影响。因此, 对于空间大尺度下研究养殖环境中氮转化相关微生物动态变化规律有待进一步开展。
4 结论(1) 缢蛏塘和肥水塘沉积物厌氧氨氧化速率范围分别为0.94~1.26和1.61~1.91 nmol/(g·h), 分别占总氮气产生量的6.09%~8.06%和8.73%~9.63%。肥水塘厌氧氨氧化速率和相对贡献显著高于缢蛏塘。
(2) 缢蛏塘和肥水塘厌氧氨氧化微生物由Ca. Scalindua、Ca. Kuenenia、Ca. Brocadia和Ca. Anammoximicrobium四个属组成。其中, Ca. Kuenenia为主要的优势属。肥水塘沉积物厌氧氨氧化微生物多样性和脱氮活性显著高于缢蛏塘(P < 0.05)。
(3) 养殖动物扰动以及频繁交换水等外界影响可能使得缢蛏塘沉积物厌氧氨氧化速率低于营养盐充足、微生物多样性复杂且沉积环境相对稳定的肥水塘。养殖塘中DO、NH4+和NO3-含量是影响厌氧氨氧化微生物脱氮的主要理化因子。
参考文献
丁维新, 袁俊吉, 刘德燕, 等, 2020. 淡水养殖系统温室气体CH4和N2O排放量研究进展[J]. 农业环境科学学报, 39(4): 749-761. |
王文卿, 张林, 张雅棉, 等, 2021. 红树林退塘还林研究进展[J]. 厦门大学学报(自然科学版), 60(2): 348-354. |
王衫允, 祝贵兵, 曲冬梅, 等, 2012. 白洋淀富营养化湖泊湿地厌氧氨氧化菌的分布及对氮循环的影响[J]. 生态学报, 32(21): 6591-6598. |
毛铁墙, 董宏坡, 陈法锦, 等, 2020. 湛江湾沉积物中反硝化和厌氧氨氧化细菌的丰度、多样性及分布特征[J]. 海洋与湖沼, 51(1): 59-74. |
石亚飞, 石善恒, 黄晓敏, 2022. 基于R的结构方程模型在生态学中的应用[J]. 生态学杂志, 41(5): 1015-1023. |
吕昊泽, 刘健, 陈锦辉, 等, 2016. 盐度对长江口3种滤食性贝类滤水率、摄食率、同化率的影响[J]. 海洋科学, 40(8): 10-17. |
朱文君, 袁晟译, 李健, 等, 2022. 菲律宾蛤仔对养殖池塘沉积物/水界面反硝化和厌氧氨氧化反应速率的影响[J]. 渔业科学进展, 43(3): 110-117. |
刘璐, 祝贵兵, 夏超, 等, 2016. 青藏高原不同海拔梯度厌氧氨氧化细菌丰度及其生物多样性空间分布[J]. 环境科学学报, 36(4): 1298-1308. |
杨雪琴, 连英丽, 颜庆云, 等, 2018. 滨海湿地生态系统微生物驱动的氮循环研究进展[J]. 微生物学报, 58(4): 633-648. |
张亦弛, 乔雪姣, 余珂, 2022. 厌氧氨氧化细菌及其群落内细菌互作关系研究进展[J]. 微生物学通报, 49(2): 737-755. |
赵永强, 夏永秋, 李博伦, 等, 2014. 利用膜进样质谱同时测定河流沉积物反硝化和厌氧氨氧化[J]. 农业环境科学学报, 33(4): 794-802. |
洪义国, 李益本, 吴佳鹏, 等, 2019. 高通量测序分析多种典型生境中厌氧氨氧化细菌的多样性分布特征[J]. 微生物学通报, 46(11): 2865-2876. |
祝贵兵, 2020. 陆地和淡水生态系统新型微生物氮循环研究进展[J]. 微生物学报, 60(9): 1972-1984. |
柴欣如, 许文军, 张东旭, 等, 2022. 缢蛏混养对三疣梭子蟹-日本囊对虾综合养殖系统理化环境和小型底栖动物的影响[J]. 海洋与湖沼, 53(3): 718-725. |
彭强, 关傲梅, 齐维晓, 等, 2021. 基于同位素示踪和分子生物学技术对潮白河沉积物中氮还原功能特征的研究[J]. 环境科学学报, 41(11): 4615-4624. |
程金, 黄文卿, 张世昌, 等, 2023. 福建省表层土壤有机碳密度空间分布及影响因素分析[J]. 农业资源与环境学报, 40(4): 805-816. |
谢成军, 宋国栋, 刘素美, 等, 2020. 自组装膜进样质谱系统及其在砂质沉积物异化硝酸盐还原研究中的应用[J]. 海洋学报, 42(2): 22-29. |
鲍士旦, 2000. 土壤农化分析[M]. 3版. 北京: 中国农业出版社.
|
ALONGI D M, PFITZNER J, TROTT L A, et al, 2005. Rapid sediment accumulation and microbial mineralization in forests of the mangrove Kandelia candel in the Jiulongjiang Estuary, China[J]. Estuarine, Coastal and Shelf Science, 63(4): 605-618. DOI:10.1016/j.ecss.2005.01.004 |
AWATA T, OSHIKI M, KINDAICHI T, et al, 2013. Physiological characterization of an anaerobic ammonium-oxidizing bacterium belonging to the "Candidatus scalindua" group[J]. Applied and Environmental Microbiology, 79(13): 4145-4148. DOI:10.1128/AEM.00056-13 |
BONAGLIA S, HYLÉN A, RATTRAY J E, et al, 2017. The fate of fixed nitrogen in marine sediments with low organic loading: an in situ study[J]. Biogeosciences, 14(2): 285-300. DOI:10.5194/bg-14-285-2017 |
BRANDSMA J, VAN DE VOSSENBERG J, RISGAARD-PETERSEN N, et al, 2011. A multi-proxy study of anaerobic ammonium oxidation in marine sediments of the Gullmar Fjord, Sweden[J]. Environmental Microbiology Reports, 3(3): 360-366. DOI:10.1111/j.1758-2229.2010.00233.x |
CAO W Z, GUAN Q S, LI Y, et al, 2017. The contribution of denitrification and anaerobic ammonium oxidation to N2 production in mangrove sediments in Southeast China[J]. Journal of Soils and Sediments, 17(6): 1767-1776. DOI:10.1007/s11368-017-1653-0 |
CHENG X J, ZENG Y X, GUO Z R, et al, 2014. Diffusion of nitrogen and phosphorus across the sediment-water interface and in seawater at aquaculture areas of Daya Bay, China[J]. International Journal of Environmental Research and Public Health, 11(2): 1557-1572. DOI:10.3390/ijerph110201557 |
DALE O R, TOBIAS C R, SONG B, 2009. Biogeographical distribution of diverse anaerobic ammonium oxidizing (anammox) bacteria in Cape Fear River Estuary[J]. Environmental Microbiology, 11(5): 1194-1207. DOI:10.1111/j.1462-2920.2008.01850.x |
DALSGAARD T, THAMDRUP B, CANFIELD D E, 2005. Anaerobic ammonium oxidation (anammox) in the marine environment[J]. Research in Microbiology, 156(4): 457-464. |
HUANG M J, LIU X, ZHOU S R, et al, 2020. Asynchrony among species and functional groups and temporal stability under perturbations: Patterns and consequences[J]. Journal of Ecology, 108(5): 2038-2046. DOI:10.1111/1365-2745.13418 |
HUMBERT S, TARNAWSKI S, FROMIN N, et al, 2010. Molecular detection of anammox bacteria in terrestrial ecosystems: distribution and diversity[J]. The ISME Journal, 4(3): 450-454. DOI:10.1038/ismej.2009.125 |
KARTAL B, RATTRAY J, VAN NIFTRIK L A, et al, 2007. Candidatus "Anammoxoglobus propionicus" a new propionate oxidizing species of anaerobic ammonium oxidizing bacteria[J]. Systematic and Applied Microbiology, 30(1): 39-49. DOI:10.1016/j.syapm.2006.03.004 |
KIM Y N, ROBINSON B, LEE K A, et al, 2017. Interactions between earthworm burrowing, growth of a leguminous shrub and nitrogen cycling in a former agricultural soil[J]. Applied Soil Ecology, 110: 79-87. DOI:10.1016/j.apsoil.2016.10.011 |
LI X F, QIAN W, HOU L J, et al, 2021. Human activity intensity controls the relative importance of denitrification and anaerobic ammonium oxidation across subtropical estuaries[J]. CATENA, 202: 105260. DOI:10.1016/j.catena.2021.105260 |
LOREAU M, NAEEM S, INCHAUSTI P, et al, 2001. Biodiversity and ecosystem functioning: current knowledge and future challenges[J]. Science, 294(5543): 804-808. DOI:10.1126/science.1064088 |
MULDER A, GRAAF A A, ROBERTSON L A, et al, 1995. Anaerobic ammonium oxidation discovered in a denitrifying fluidized bed reactor[J]. FEMS Microbiology Ecology, 16(3): 177-184. DOI:10.1111/j.1574-6941.1995.tb00281.x |
QUAN Z X, RHEE S K, ZUO J E, et al, 2008. Diversity of ammonium-oxidizing bacteria in a granular sludge anaerobic ammonium-oxidizing (anammox) reactor[J]. Environmental Microbiology, 10(11): 3130-3139. DOI:10.1111/j.1462-2920.2008.01642.x |
RISGAARD-PETERSEN N, MEYER R, SCHMID M, et al, 2004. Anaerobic ammonium oxidation in an estuarine sediment[J]. Aquatic Microbial Ecology, 36(3): 293-304. DOI:10.3354/ame036293 |
SCHMID M C, MAAS B, DAPENA A, et al, 2005. Biomarkers for in situ detection of anaerobic ammonium-oxidizing (anammox) bacteria[J]. Applied and Environmental Microbiology, 71(4): 1677-1684. DOI:10.1128/AEM.71.4.1677-1684.2005 |
SHAN J, ZHAO X, SHENG R, et al, 2016. Dissimilatory nitrate reduction processes in typical Chinese paddy soils: rates, relative contributions, and influencing factors[J]. Environmental Science & Technology, 50(18): 9972-9980. DOI:10.1021/acs.est.6b01765 |
SHEN L D, CHENG H X, LIU X, et al, 2017. Potential role of anammox in nitrogen removal in a freshwater reservoir, Jiulonghu Reservoir (China)[J]. Environmental Science and Pollution Research, 24(4): 3890-3899. DOI:10.1007/s11356-016-8126-7 |
SHEN L, LIU S, HUANG Q, et al, 2014. Evidence for the cooccurrence of nitrite-dependent anaerobic ammonium and methane oxidation processes in a flooded paddy field[J]. Applied and Environmental Microbiology, 80(24): 7611-7619. DOI:10.1128/AEM.02379-14 |
SHEN L D, WU H S, GAO Z Q, et al, 2016. Evidence for anaerobic ammonium oxidation process in freshwater sediments of aquaculture ponds[J]. Environmental Science and Pollution Research, 23(2): 1344-1352. DOI:10.1007/s11356-015-5356-z |
SONTHIPHAND P, HALL M W, NEUFELD J D, 2014. Biogeography of anaerobic ammonia-oxidizing (anammox) bacteria[J]. Frontiers in Microbiology, 5: 399. DOI:10.3389/fmicb.2014.00399 |
SUENAGA T, OTA T, OBA K, et al, 2021. Combination of 15N tracer and microbial analyses discloses N2O sink potential of the anammox community[J]. Environmental Science & Technology, 55(13): 9231-9242. DOI:10.1021/acs.est.1c00674 |
TEIXEIRA C, MAGALHÃES C, JOYE S B, et al, 2016. Response of anaerobic ammonium oxidation to inorganic nitrogen fluctuations in temperate estuarine sediments[J]. Journal of Geophysical Research: Biogeosciences, 121(7): 1829-1839. DOI:10.1002/2015jg003287 |
THAMDRUP B, DALSGAARD T, 2002. Production of N2 through anaerobic ammonium oxidation coupled to nitrate reduction in marine sediments[J]. Applied and Environmental Microbiology, 68(3): 1312-1318. DOI:10.1128/AEM.68.3.1312-1318.2002 |
WANG S Y, ZHU G B, PENG Y Z, et al, 2012. Anammox bacterial abundance, activity, and contribution in riparian sediments of the Pearl River estuary[J]. Environmental Science & Technology, 46(16): 8834-8842. DOI:10.1021/es3017446 |
WELLS N S, MAHER D T, ERLER D V, et al, 2018. Estuaries as sources and sinks of N2O across a land use gradient in Subtropical Australia[J]. Global Biogeochemical Cycles, 32(5): 877-894. DOI:10.1029/2017gb005826 |
WU J P, HONG Y G, LIU X H, et al, 2021. Variations in nitrogen removal rates and microbial communities over sediment depth in Daya Bay, China[J]. Environmental Pollution, 286: 117267. DOI:10.1016/j.envpol.2021.117267 |
WU R S S, LAM K S, MACKAY D W, et al, 1994. Impact of marine fish farming on water quality and bottom sediment: A case study in the sub-tropical environment[J]. Marine Environmental Research, 38(2): 115-145. DOI:10.1016/0141-1136(94)90004-3 |
YA T, DU S, LI Z Y, et al, 2021. Successional dynamics of molecular ecological network of anammox microbial communities under elevated salinity[J]. Water Research, 188: 116540. DOI:10.1016/j.watres.2020.116540 |
YANG P, ZHANG Y F, LAI D Y F, et al, 2018. Fluxes of carbon dioxide and methane across the water-atmosphere interface of aquaculture shrimp ponds in two subtropical estuaries: The effect of temperature, substrate, salinity and nitrate[J]. Science of The Total Environment, 635: 1025-1035. DOI:10.1016/j.scitotenv.2018.04.102 |
ZHANG Y P, RUAN X H, WAN Y, et al, 2016. Effects of environmental factors on anammox bacterial community structure in sediments of a freshwater aquaculture farm, Yangcheng Lake[J]. Geomicrobiology Journal, 33(6): 479-487. DOI:10.1080/01490451.2015.1054005 |
ZHENG Y L, JIANG X F, HOU L J, et al, 2016. Shifts in the community structure and activity of anaerobic ammonium oxidation bacteria along an estuarine salinity gradient[J]. Journal of Geophysical Research: Biogeosciences, 121(6): 1632-1645. DOI:10.1002/2015jg003300 |
ZHU G B, WANG S Y, WANG Y, et al, 2011. Anaerobic ammonia oxidation in a fertilized paddy soil[J]. The ISME Journal, 5(12): 1905-1912. DOI:10.1038/ismej.2011.63 |