 2023, Vol. 54
2023, Vol. 54

中华绒螯蟹“长荡湖1号”新品系选育第二代(G2) 生长、性腺发育和养殖性能的研究

http://dx.doi.org/10.11693/hyhz20230200032
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 张光宝, 姜晓东, 庄振俊, 陈文彬, 罗明, 成永旭, 陈晓武, 吴旭干. 2023.
- ZHANG Guang-Bao, JIANG Xiao-Dong, ZHUANG Zhen-Jun, CHEN Wen-Bin, LUO Ming, CHENG Yong-Xu, CHEN Xiao-Wu, WU Xu-Gan. 2023.
- 中华绒螯蟹“长荡湖1号”新品系选育第二代(G2) 生长、性腺发育和养殖性能的研究
- EVALUATION ON GROWTH, GONADAL DEVELOPMENT, AND CULTURE PERFORMANCE OF "CHANGDANG LAKE 1" NEW STAIN OF SECOND-GENERATION OF CHINESE MITTEN CRAB (ERIOCHEIR SINENSIS)
- 海洋与湖沼, 54(5): 1444-1453
- Oceanologia et Limnologia Sinica, 54(5): 1444-1453.
- http://dx.doi.org/10.11693/hyhz20230200032
文章历史
-
收稿日期:2023-02-14
收修改稿日期:2023-05-10
引用本文
张光宝, 姜晓东, 庄振俊, 陈文彬, 罗明, 成永旭, 陈晓武, 吴旭干. 2023. 中华绒螯蟹“长荡湖1号”新品系选育第二代(G2) 生长、性腺发育和养殖性能的研究[J]. 海洋与湖沼, 54(5): 1444-1453

ZHANG Guang-Bao, JIANG Xiao-Dong, ZHUANG Zhen-Jun, CHEN Wen-Bin, LUO Ming, CHENG Yong-Xu, CHEN Xiao-Wu, WU Xu-Gan. 2023. EVALUATION ON GROWTH, GONADAL DEVELOPMENT, AND CULTURE PERFORMANCE OF "CHANGDANG LAKE 1" NEW STAIN OF SECOND-GENERATION OF CHINESE MITTEN CRAB (ERIOCHEIR SINENSIS)[J]. Oceanologia et Limnologia Sinica, 54(5): 1444-1453.
中华绒螯蟹“长荡湖1号”新品系选育第二代(G2) 生长、性腺发育和养殖性能的研究
1. 上海海洋大学 农业农村部淡水种质资源重点实验室 上海 201306;
2. 常州市金坛区水产技术推广中心 江苏常州 213200;
3. 上海市水产动物良种创制与绿色养殖协同创新中心 上海 201306;
4. 上海海洋大学 水产科学国家级实验教学示范中心 上海 201306
2. 常州市金坛区水产技术推广中心 江苏常州 213200;
3. 上海市水产动物良种创制与绿色养殖协同创新中心 上海 201306;
4. 上海海洋大学 水产科学国家级实验教学示范中心 上海 201306
摘要:“长荡湖1号”A系和B系是分别以成活率和大规格为主要选育指标建立的两个相互独立的中华绒螯蟹(Eriocheir sinensis)选育品系, 为评估“长荡湖1号”偶数年群体(偶数年繁殖孵化)选育第二代(G2)的选育效果, 本实验以未选育群体作为对照组, 比较了两选育品系和对照组在实际池塘养殖条件下的生长、性腺发育和养殖性能。结果显示: (1) 两选育品系A系和B系在扣蟹、成蟹养殖阶段的最终平均体质量均高于对照组, 其中B系雌体与对照组具有显著差异(P < 0.05); 无论雌体还是雄体, B系最终平均体质量均略高于A系, 但无显著差异(P > 0.05); (2) 各组实验蟹雌雄个体在成蟹阶段的生殖蜕壳高峰期分别为2021年8月10~30日和8月30日~9月20日, 各组成蟹完成生殖蜕壳后性腺迅速发育, 并且性腺发育后期, 对照组GSI最高、B系的GSI最低; (3) 就成活率而言, 三组蟹在扣蟹和成蟹阶段的成活率大小顺序为: A系 > 对照组 > B系, 差异不显著(P > 0.05); 而在产量、大规格蟹比例和饲料系数方面, 两选育品系均优于对照组。综上, 经过两代配套系选育, “长荡湖1号”A系和B系均获得良好的选育进展, 其中A系具有高成活率和高产量的特点, B系具有平均体质量大、大规格成蟹比例高和饲料系数低等优势。
关键词:中华绒螯蟹 配套系选育 池塘养殖 养殖性能 遗传进展
EVALUATION ON GROWTH, GONADAL DEVELOPMENT, AND CULTURE PERFORMANCE OF "CHANGDANG LAKE 1" NEW STAIN OF SECOND-GENERATION OF CHINESE MITTEN CRAB (ERIOCHEIR SINENSIS)
1. Key Laboratory of Freshwater Germplasm Resources, Ministry of Agriculture and Rural Affairs, Shanghai Ocean University, Shanghai 201306, China;
2. Changzhou Jintan District Fisheries Technical Promotion Center, Changzhou 213200, China;
3. Shanghai Collaborative Innovation Center for Cultivating Elite Breeds and Green-culture of Aquaculture animals, Shanghai 201306, China;
4. National Demonstration Centre for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China
2. Changzhou Jintan District Fisheries Technical Promotion Center, Changzhou 213200, China;
3. Shanghai Collaborative Innovation Center for Cultivating Elite Breeds and Green-culture of Aquaculture animals, Shanghai 201306, China;
4. National Demonstration Centre for Experimental Fisheries Science Education, Shanghai Ocean University, Shanghai 201306, China
Abstract: The "Changdang Lake 1" Strains A and B of Chinese mitten crab (Eriocheir sinensis) were selected in terms of the economic indices of survival and growth performance, respectively. The growth, gonadal development, and culture performance of the second-generation (G2) of the "Changdang Lake 1" were evaluated in comparison with the control. Results show that the average body weight of two selected strains were greater than that of the control in the juvenile and adult culture stages, and the final body weight of females in Strain B was significantly greater than the control (P < 0.05). In addition, the average body weight of crabs in Strain B was always higher than Strain A during the culture experiments with no significant difference (P > 0.05). The puberty molting of female and male in all groups concentrated on Aug.10~30 and Aug.30~Sep.20, respectively. The gonadal development initiated soon after completing the puberty molting. In late gonad development, the maximum gonadosomatic index (GSI) occurred in the control group and the minimum in Strain B group. The survival rate of Strain A in the juvenile and adult stages was 6.77% and 7.76% higher than the control group, respectively, but the survival rate of Strain B was slightly smaller than the control. The final yield and the percentage of large-sized individuals in the two strains were greater than that of the control, while the feed conversion rate (FCR) was relatively smaller in the two selected strains. Therefore, the two crab strains achieved expected economic values after two generations of selection, of which Strain A showed greater survival rate and yield, and Strain B presented more large-sized individuals, larger adult size, and lower FCR.
Key words:
Eriocheir sinensis complete-set line breeding pond culture culture performance genetic gain
中华绒螯蟹(Eriocheir sinensis)俗称河蟹、大闸蟹, 是我国重要的养殖蟹类之一(堵南山, 1998), 其因味道鲜美、营养丰富而广受消费者喜爱(Wang et al, 2016; Long et al, 2019)。进入21世纪之后, 中华绒螯蟹野生资源有所恢复, 但如今市场上的商品蟹仍以养殖为主, 连续多年养殖产量均在80万t左右(吴晓峰等, 2020; 农业农村部渔业渔政管理局等, 2021)。目前我国中华绒螯蟹产业仍存在一些问题, 如盲目引种、亲本规格小和近亲繁殖等, 造成部分中华绒螯蟹群体种质退化, 商品成蟹品质参差不齐(张世勇等, 2013)。选择育种是常用的传统有效的遗传改良方法之一, 通过对群体的反复选择淘汰、优中选优, 从而育成符合选育目标的新品种(邹杰等, 2013)。该方法能够分离并固定优良性状, 保留有益的变异, 提高经济性状, 提纯和复壮中华绒螯蟹品种(朱华平等, 2010)。中国在中华绒螯蟹良种选育工作上取得一定的成果, 迄今为止已培育出五个中华绒螯蟹养殖新品种(Wang et al, 2018b), 但这些新品种仅以单一的生长性能作为指标进行选育, 尚不能满足中华绒螯蟹多样化的市场需求(Wang et al, 2018a)。
长荡湖是长江中下游地区的天然浅水湖泊, 湖底平坦, 天然饵料丰富, 其湖岸周边中华绒螯蟹养殖规模不断扩大, 已逐渐成为江苏金坛农业中的特色产业。但近年来长荡湖地区蟹种质退化严重, 出现商品成蟹规格较小、成活率下降和养殖效果不稳定等问题, 极大阻碍了长荡湖地区中华绒螯蟹产业健康发展和升级。为选育具有长荡湖地区特性的优良中华绒螯蟹新品系, 上海海洋大学和江苏金坛区水产技术推广中心自2017年起开展合作, 分别以“雌体≥200 g, 雄体≥10 g”和“雌体≥250 g, 雄体≥360 g”这两种规格的长荡湖地区优质亲本作为奠基群体进行“长荡湖1号”A系和B系的中华绒螯蟹新品系选育, 其中“长荡湖1号”A系以高成活率为选育目标, “长荡湖1号”B系以大规格作为选育目标(庄振俊等, 2022), 目前已经选育到第三代(G3)扣蟹阶段。罗明等(2020)和庄振俊等(2021)分别对“长荡湖1号”G1在扣蟹和成蟹养殖阶段的生长性能和养殖效果进行评估, 结果表明在整个养殖阶段两选育品系子一代相较于未选育群体, 均表现出一定的优良特性, 在成蟹养殖阶段, A系成活率和平均体质量分别提高了22.9%和11.7%, B系成活率和平均体质量分别提高了5.5%和21.9%。
为进一步评估两选育品系目标性状在G2阶段的最新选育进展, 本实验将两选育品系大眼幼体在相似的网箱环境下从仔蟹养至扣蟹, 随后挑选规格一致的两品系扣蟹在相似池塘条件下养殖至成蟹, 并以未选育群体作为对照组, 分别评估“长荡湖1号”偶数年G2在扣蟹和成蟹阶段的生长、性腺发育及养殖性能, 用于进一步开展中华绒螯蟹的种质资源评价和良种培育研究, 以期为新品种进一步推广应用提供理论依据。
1 材料与方法 1.1 亲本来源和选育过程实验用“长荡湖1号”A系、B系和对照组的奠基群均源自金坛区成蟹养殖池塘。中华绒螯蟹的生命周期通常为两周年, 性成熟蟹只繁殖一代, 不能进行多年份重复繁殖, 因此, 同一地区相邻两个年份的中华绒螯蟹群体不存在基因交流, 而分为奇数年群体(奇数年孵化)和偶数年群体(偶数年孵化)。本实验蟹亲本于2017年底挑选, 2018年4~6月繁殖孵化出大眼幼体进行一龄幼蟹培育, 2019年11月挑选成蟹作为G2亲本(表 1), 2020年5月在上海海洋大学崇明产学研基地开展扣蟹阶段养殖, 次年3月在金坛区水产站指前基地进行成蟹养殖实验, 目前已选育至偶数年第二代(G2)成蟹阶段。“长荡湖1号”的偶数年群体的选育过程示意图见图 1。
表 1 中华绒螯蟹“长荡湖1号b偶数年子二代(G2)的亲本信息 Tab. 1 Broodstocks information for the second selective generation (G2) of even-year E. sinensis "Changdang Lake 1"
| 品系 | 雌体 | 雄体 | |||
| 数量 | 平均体质量/g | 数量 | 平均体质量/g | ||
| 长荡湖1号A系 | 1 018 (≥200 g) | 210.93±8.24 | 387 (≥300 g) | 321.95±14.58 | |
| 长荡湖1号B系 | 526 (≥250 g) | 262.92±7.09 | 225 (≥355 g) | 378.41±15.37 | |
| 对照组 | 252 (100~160 g) | 125.81±7.82 | 103 (150~210 g) | 168.53±11.25 | |
 |
| 图 1 中华绒螯蟹“长荡湖1号”的偶数年群体的选育过程示意图 Fig. 1 Schematic diagram of the breeding course of even-year population of E. sinensis "Changdang Lake 1" |
实验设置12个面积相等的围隔(长×宽=7.8 m×7.8 m)作为养殖池塘用于比较两选育品系和对照组在扣蟹阶段的养殖性能差异。围隔中间各有1个水坑(长×宽×深=6 m×4 m×0.7 m), 在水坑中设置1个网箱(长×宽×高=2 m×2 m×1 m), 扣蟹阶段养殖实验在网箱中进行。各网箱中大眼幼体投放密度约为375只/m2, 每实验组设置4个平行网箱。网箱内放置伊乐藻(Elodea nuttallii)供幼蟹隐蔽和蜕壳, 水坑四周土地种植水稻以净化水质。
由于扣蟹前期蜕壳频率高, 成活率不稳定, 导致各网箱蟹养殖效果差距悬殊。因此在2020年6月中旬各网箱内伊乐藻清除后, 重新调整豆蟹数量(400只, 雌雄各半), 并放置适量水花生(Alternanthera philoxeroides)。配合饲料在每天下午5:00投喂, 大眼幼体至豆蟹阶段日投喂破碎料量为1~5 g, 随着扣蟹的蜕壳和生长, 依次改用1.2、1.6和1.8 mm粒径的颗粒饲料, 每日投饲量为幼蟹总体质量的1%~6%, 根据气候和摄食情况进行适当调整。
1.3 成蟹实验池塘和日常管理2021年3月初挑选整齐、有活力、附肢健全的扣蟹, 聚维酮碘溶液浸泡消毒后用于成蟹阶段养殖实验。选用9口面积相同(长×宽=23 m×23 m)的室外池塘, 每个实验塘放养扣蟹1 200只, 雌雄各半。
扣蟹放养前用含氯石灰对实验池塘进行清塘。二周后在池塘底部种植伊乐藻, 养殖后期再补种轮叶黑藻(Hydrilla verticillata)和少量水花生。5月初在每个池塘中投放约100 kg螺蛳(Margarya melanioides)作为补充饵料以及净化水质, 投放5尾鳜(Siniperca chuatsi)用于控制野生杂鱼数量。成蟹配合饲料(浙江澳华饲料公司生产)投喂时间为每天下午5:00, 投喂量为蟹体质量的1.0%~3.0%。使用精创RC-4温度记录仪记录水温, 外置探头放置深度为水下20 cm, 每15 min记录一次水温。池塘底部铺设管道增氧设施, 在阴雨天及高温夜间对池塘水体进行微孔曝气增氧。定期使用水质测试盒(厦门迈凯威生物科技有限公司生产)检测每个成蟹塘的氨氮(< 0.4 mg/L)、溶氧(> 3 mg/L)、pH (7.0~9.0)以及亚硝酸盐(< 0.15 mg/L)等水质指标。每天早晚各巡塘一次, 观察水体颜色和水位, 为改善池塘水质, 每二周更换掉池塘内10%~ 30%的水体。
1.4 数据采集 1.4.1 扣蟹养殖阶段生长性能2020年6月中旬将各网箱里所有的豆蟹捞出, 分别统计每个网箱内豆蟹数量, 使用电子天平称总体质量(精确到0.01 g)后, 分别计算平均体质量、成活率(survival rate, SR, %)以及增重率(weight growth rate, WGR, %)。调整密度后, 每月15日从每个网箱里随机采样雌雄体各100只, 称质量后计算每月的增重率及特定生长率(specific growth rate, SGR, %/d), 计算公式如下:
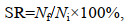 (1)
(1)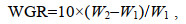 (2)
(2)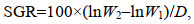 (3)
(3)式中, Nf为成活个体数量; Ni为最初投放苗种数量; W1和W2分别为日龄1和2时各组蟹的平均体质量(单位: g), D为采样的间隔天数(单位: d)。
1.4.2 扣蟹养殖阶段养殖性能扣蟹养殖实验结束后, 统计各网箱内存活的扣蟹数量并称体质量, 分别计算成活率、产量(yield, Y, g/m2)和饲料系数(feed conversion ratio, FCR)。按体质量对扣蟹进行规格分级, 雌雄蟹均分为4级: < 4.00 g、4.00~5.99 g、6.00~ 7.99 g和≥8.00 g, 统计两选育品系和对照组扣蟹各规格等级所占的比例。
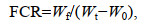 (4)
(4)式中, Wf为总消耗饲料量(g); Wt为扣蟹总质量(g); W0为投放苗种总质量(g)。
1.4.3 成蟹养殖阶段生长性能和性腺发育根据中华绒螯蟹在成蟹阶段的蜕壳规律, 分别于2021年4、5、6、8和10月中旬从每个池塘随机采集雌雄个体各20只, 称量蟹体质量后计算WGR和SGR。为观察各池塘蟹生殖蜕壳情况, 7月20日起每隔20 d从各养殖池塘随机采样40只个体, 雌雄各半, 统计已完成生殖蜕壳的雌雄个体数量, 计算生殖蜕壳率(puberty molting rate, %)。采样个体是否已经完成生殖蜕壳采用王武等(2013)的方法进行判断。
当生殖蜕壳率达到80%后, 分别从9月10日和9月20日起每隔30 d从各实验池塘随机采集3只雌、雄蟹, 称量体质量后, 解剖得到肝胰腺和性腺并称质量。性腺指数(gonadosomatic index, GSI)和肝胰腺指数(hepatosomatic index, HSI)的计算公式如下:
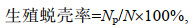 (5)
(5)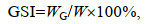 (6)
(6)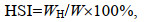 (7)
(7)式中, Np为已完成生殖蜕壳的蟹数量(单位: 只); N为采样蟹总数(单位: 只); WG为蟹性腺质量(单位: g); WH为蟹肝胰腺质量(单位: g); W为蟹体质量(单位: g)。
1.4.4 成蟹养殖阶段成活率、产量、饲料系数及规格分布2021年11月25日左右, 通过地笼将大部分商品蟹起捕后, 排干池水捡蟹清塘, 分别统计各池塘雌雄个体成活率、平均体质量、产量及饲料系数。参照庄振俊等(2021)的方法对捕获成蟹进行规格分级, 记录各体质量等级成蟹数量。
1.5 数据分析实验数据分析采用SPSS 26.0软件, 均以平均值±标准差(Mean±SD)表示。两选育品系和对照组之间的差异分析采用one-way ANOVA法, 并采用Duncan氏法进行多重比较, 取P < 0.05为差异显著。
2 结果 2.1 扣蟹阶段的养殖性能 2.1.1 大眼幼体至仔蟹期间养殖性能3组仔蟹的平均体质量大小顺序为B系 > A系 > 对照组(表 2), 各组间无显著差异(P > 0.05), 均在0.45 g左右。3组仔蟹增重率的变化情况与体质量相类似。就成活率而言, 对照组成活率比B系高6%左右, 但差异不显著(P > 0.05)。
表 2 中华绒螯蟹“长荡湖1号”偶数年G2在大眼幼体至仔蟹阶段养殖性能 Tab. 2 Culture performance of the second generation (G2) of E. sinensis in the juvenile stage
| 项目 | 长荡湖1号A系 | 长荡湖1号B系 | 对照组 |
| 平均体质量/g | 0.45±0.01 | 0.48±0.04 | 0.43±0.02 |
| 增重率/% | 66.97±1.81 | 71.93±5.97 | 64.43±2.50 |
| 成活率/% | 59.51±4.05 | 56.36±5.45 | 62.69±3.08 |
两选育品系扣蟹在2020年8~11月份的平均体质量高于对照组(图 2a, 2d), 其中B系雌体体质量在7、8及11月份的平均体质量显著高于对照组(P < 0.05)。随着养殖实验的进行, 三组扣蟹的WGR整体表现出下降趋势(图 2b, 2e), 但各组扣蟹之间WGR均无显著差异。两选育品系和对照组扣蟹SGR的变化趋势与WGR类似(图 2c, 2f)。
 |
| 图 2 中华绒螯蟹“长荡湖1号”偶数年G2在扣蟹阶段生长性能比较 Fig. 2 Comparison in growth performance of "Changdang Lake 1" even-year G2 in the juvenile stage 注: a. 雄体平均体质量; b. 雄体增重率; c. 雄体特定生长率; d. 雌体平均体质量; e. 雌体增重率; f. 雌体特定生长率 |
三组扣蟹最终平均体质量的大小次序为B系 > A系 > 对照组(表 3), 其中B系雌体与对照组相比有显著差异(P < 0.05)。在成活率方面, 不论雌体还是雄体, 长荡湖A系均最高, B系雄体最低, 整体而言, B系成活率略低于对照组, 但各组之间显著不差异(P > 0.05)。两选育品系的最终产量比对照组高20%左右, 但两选育品系最终产量较为接近, 均为300 g/m2左右。在饲料系数方面, 两选育品系比对照组低12%左右, 而两选育品系饲料系数较为接近, 均为1.85左右。
表 3 中华绒螯蟹“长荡湖1号”偶数年G2扣蟹平均体质量、成活率、产量和饲料系数 Tab. 3 Average body mass, survival, yield, and feed conversion rate of "Changdang Lake 1" even-year G2 in the juvenile stage
| 项目 | A系 | B系 | 对照 |
| 平均体质量/g | |||
| 雄体 | 6.30±1.11 | 6.59±0.93 | 5.90±0.63 |
| 雌体 | 5.94±0.09ab | 6.54±0.46b | 5.12±0.55a |
| 整体 | 6.12±0.59 | 6.57±0.69 | 5.51±0.59 |
| 成活率/% | |||
| 雄体 | 49.50±3.54 | 42.50±8.20 | 45.00±4.71 |
| 雌体 | 50.17±4.40 | 49.33±9.45 | 48.33±7.79 |
| 整体 | 49.83±3.57 | 45.92±8.80 | 46.67±6.25 |
| 产量/(g/m2) | |||
| 雄体 | 157.99±40.13 | 136.43±13.88 | 131.63±10.82 |
| 雌体 | 148.95±13.31 | 159.34±21.45 | 121.95±12.89 |
| 整体 | 306.85±50.81 | 295.67±33.19 | 254.21±23.35 |
| 饲料系数 | 1.85±0.51 | 1.86±0.14 | 2.11±0.15 |
| 注: 同行数据中上标有不同字母代表差异显著(P < 0.05)。下同 | |||
三组扣蟹各规格所占百分比均呈正态分布, 且主要分布于4.00~5.99 g和6.00~7.99 g这两个规格区间内, 占总体60%以上(图 3)。无论雄体还是雌体, 对照组扣蟹在 < 4.00 g规格范围内所占的比例均高于A系和B系, 其中雌体差异显著(P < 0.05)。此外, B系扣蟹在≥8.00 g规格范围内所占的比例均高于A系和对照组扣蟹, 其中B系雌体所占比例显著高于对照组(P < 0.05)。
 |
| 图 3 中华绒螯蟹“长荡湖1号”偶数年G2在扣蟹阶段规格分布的情况 Fig. 3 Harvest size distribution of different body mass ranges of the "Changdang Lake 1" in even-year G2 in the juvenile stage 注: a. 扣蟹雄体的规格分布; b. 扣蟹雌体的规格分布 |
如图 4所示, B系在2021年5月份后表现出生长优势(图 4a, 4d), 并且在5、8及10月份时B系雌体的平均体质量显著高于对照组(P < 0.05)。A系个体平均体质量略高于对照组, 其中5月份时雌体差异显著(P < 0.05)。两选育品系和对照组在成蟹养殖阶段的WGR整体呈下降趋势(图 4b, 4e), 其中B系在4~5月及6~8月期间的增重率略高于其余两组, 但无显著差异(P > 0.05)。两选育品系和对照组中华绒螯蟹SGR变化情况与WGR相似(图 4c, 4e), 各组蟹在养殖前期(3~6月)的SGR可达2%左右, 但8月份之后雄蟹SGR小于1%, 雌蟹SGR小于0.8%。
 |
| 图 4 “长荡湖1号”偶数年G2成蟹养殖阶段的生长比较 Fig. 4 Comparison of the growth performance of "Changdang Lake 1" in even-year G2 in the adult stage 注: a. 雄体平均体质量; b. 雄体增重率; c. 雄体特定生长率; d. 雌体平均体质量; e. 雌体增重率; f. 雌体特定生长率 |
三组蟹中雌体的生殖蜕壳高峰期位于2021年8月10~30日期间(图 5a)、雄体生殖蜕壳高峰期位于8月30日~9月20日期间(图 5d), 但三组中华绒螯蟹生殖蜕壳率在7~10月均无显著差异(P > 0.05)。各组实验蟹GSI在性腺发育期间呈升高趋势, 其中10月10日时B系雌体GSI显著高于其余两组(P < 0.05), 但性腺发育后期对照组GSI最高而B系GSI最低。各组实验蟹雌体HSI在9~11月呈下降趋势, 其中雌体HSI在9月10日高于10%, 而11月10日降至7%左右; 两选育品系雄体HSI在9~11月先略微升高后降低, 对照组雄体HSI逐月下降, 其中雄体HSI在9月20日为8%左右, 到11月20日降至6.5%左右。
 |
| 图 5 “长荡湖1号”偶数年G2生殖蜕壳率、性腺指数和肝胰腺指数的比较 Fig. 5 Comparison of puberty molting rate, GSI and HSI of "Changdang Lake 1" in even-year G2 in the adult stage 注: a. 雄体生殖蜕壳率; b. 雄体性腺指数; c. 雄体肝胰腺指数; d. 雌体生殖蜕壳率; e. 雌体性腺指数; f. 雌体肝胰腺指数 |
两选育品系在成蟹阶段的最终平均体质量均高于对照组(表 4), 其中B系雌体体质量及雌雄整体体质量显著高于对照组(P < 0.05)。成活率方面, A系最终雌雄整体成活率最高, B系整体成活率最低, 对照组成活率位于A系和B系之间, 但各组之间无显著差异。产量方面, A系最终雌雄整体产量最高, 对照组整体产量最低(P > 0.05)。此外, 各组饲料系数的大小顺序为: 对照组 > A系 > B系。
表 4 “长荡湖1号”偶数年G2成蟹养殖阶段最终平均体质量、成活率、产量和饲料系数比较 Tab. 4 Average body mass, survival, yield and feed conversion ratio of "Changdang Lake 1" in even-year G2 in the adult stage
| 项目 | A系 | B系 | 对照 |
| 平均体质量/g | |||
| 雄体 | 205.15±27.55 | 215.86±17.95 | 180.32±25.70 |
| 雌体 | 150.83±18.12ab | 172.53±4.43a | 135.00±13.22b |
| 整体 | 175.72±21.73ab | 192.59±10.77a | 154.08±18.57b |
| 成活率/% | |||
| 雄体 | 35.96±6.49 | 30.50±3.61 | 30.83±9.33 |
| 雌体 | 42.83±9.91 | 35.39±1.95 | 42.28±11.87 |
| 整体 | 39.40±7.95 | 32.94±2.48 | 36.56±10.57 |
| 产量/(g/m2) | |||
| 雄体 | 81.82±10.81 | 73.99±10.72 | 61.04±11.72 |
| 雌体 | 72.22±16.42 | 68.64±3.62 | 63.31±14.07 |
| 整体 | 154.04±26.60 | 142.63±12.68 | 124.35±25.61 |
| 饲料系数 | 2.73±0.86 | 2.42±0.40 | 3.14±0.60 |
各组雄蟹体质量主要集中于150~249.9 g规格区间内(图 6), B系在大规格(≥225 g)分布比例方面明显高于A系和对照组, 其中B系个体在225~249.9 g和250~274.9 g规格范围内的比例显著高于对照组个体(P < 0.05)。而B系在小规格(< 174.9 g)比例方面明显低于其余两组, 并且B系个体在125~149.9 g和150~174.9 g规格范围内的比例显著低于对照组个体(P < 0.05)。各组雌蟹体质量主要集中于125~174.9 g规格区间内, 其中B系个体在175~199.9 g规格范围内的比例显著高于对照组个体(P < 0.05)。
 |
| 图 6 “长荡湖1号”偶数年G2养殖成蟹的规格分布比较 Fig. 6 Harvest size distribution of different body mass ranges of the "Changdang Lake 1" in even-year G2 in the adult stage 注: a. 成蟹雄体的规格分布; b. 成蟹雌体的规格分布 |
甲壳动物生长性能相关的经济性状均具有较高的遗传力, 因此科研工作者经常通过遗传选育的手段提升甲壳动物生长性能(Preston et al, 2004)。本实验中两选育品系在扣蟹阶段的最终平均体质量分别比对照组高11.07%和19.23%, 在成蟹阶段的最终平均体质量分别比对照组高14.04%和24.99%, 并且B系个体在扣蟹和成蟹阶段的最终平均体质量分别比A系高7.35%和9.60%, 这些结果均说明使用大规格中华绒螯蟹作为亲本可以有效提升子代生长性能, 同时也验证了中华绒螯蟹生长性状的高遗传力。大规格中华绒螯蟹亲本在育种方面的优势之前已有诸多研究, 但主要集中于亲本规格对子代在扣蟹养殖阶段生长性能的影响(陈军伟等, 2016; 范陈伟等, 2021; 杨帅帅等, 2021), 成蟹阶段的相关研究较少。本实验结果表明大规格中华绒螯蟹亲本同样可以有效提高子代在成蟹阶段的生长性能。就不同性别个体之间的选育效果差异而言, 无论扣蟹养殖阶段还是成蟹养殖阶段, 本实验中均为雌体最终平均体质量差异显著, 具体表现为B系最终平均体质量显著高于对照组, 而两品系雄体最终体质量虽表现出一定的选育效果, 但组间并无显著差异, 凸显出本实验中雌体生长性能的选育效果好于雄体。该结果与“长荡湖1号”两品系G1在成蟹阶段选育效果评估的结果相一致, 即同样为雌体最终平均体质量差异显著(庄振俊等, 2021)。在之前鱼类生长性状遗传力评估的相关研究中也表现出类似结果, 比如在食蚊鱼(Gambusia affinis)中的研究表明雌体体质量的遗传力(Heritability, h2)为0.77, 大约为雄体(0.25)的三倍(Busack et al, 1983)。此外, 在斑点鲶鱼(Lctalurus punctatus)的研究中, 雌体生长性状的遗传力为0.52, 几乎为雄体(0.27)的两倍(El-Ibiary et al, 1978)。针对本实验中雄体生长性能选育效果较差的问题, 有必要尝试从亲本规格、亲本配对比例和选择强度等方面调整新品系选育策略, 从而改善雄体生长性能的选育效果。
处于性腺发育初期的中华绒螯蟹完成生殖蜕壳后, 随即进入性腺快速发育阶段, 在此期间蟹规格不再大幅增加, 不断储存营养物质, 为后续越冬及生殖作准备(Long et al, 2020)。本实验中各组实验蟹雌雄个体GSI均随着时间的延长逐渐上升而HSI整体呈下降趋势, 暗示中华绒螯蟹在性腺发育期间, 储存在肝胰腺中的营养物质会部分转移到性腺中。就组间差异而言, 本实验中B系个体在性腺发育后期的GSI低于对照组, 并且在之前的G1选育阶段B系个体同样表现为较慢的生殖蜕壳速度和较低的GSI (庄振俊等, 2021), B系个体在选育过程中所表现出的这种晚熟性状, 验证了以生殖蜕壳时间和性腺发育速度为指标进行选育的可行性。本研究中B系个体的生长性能与性腺发育速度呈明显的负相关, 表明以生长性能作为指标开展选择育种可能会推迟甲壳动物的性腺发育, 这可能是由于大规格甲壳动物将更多能量优先用于生长, 而用于性腺发育和繁殖的能量较为滞后(Argue et al, 2002)。虽然本实验中各组HSI在性腺发育期间整体呈下降趋势, 但B系雄体的HSI在9~11月份期间呈先增后降的变化趋势, 这可能是由于9月20日~10月20日期间A系及对照组肝胰腺中营养物质已逐渐向性腺中转移, 而B系由于成熟时间和性腺发育较晚, 性腺中营养物质的积累也相应较晚(Wu et al, 2020)。性腺和肝胰腺是中华绒螯蟹重要的可食组织, 因此本实验中遗传选育对GSI和HSI的影响可以为中华绒螯蟹组织系数的选育提供一些参考依据, 但整体上A系和B系选育组和未选育组中华绒螯蟹GSI和HSI相近, 可能环境因素起到了主要的作用, 选育效果并不明显。
成活率是影响中华绒螯蟹养殖效果和经济效益的主要因素, 也是评价其遗传育种的重要指标(林永淮等, 2009)。由于有限的生存空间和饵料资源, 传统养殖模式下商品蟹规格和成活率相互矛盾, 生长性能较好的群体往往成活率较低(董江水等, 2007)。本研究中A系在扣蟹和成蟹阶段的整体成活率均高于未选育群体, 说明以优质中华绒螯蟹作为亲本进行选育可以在一定程度上提升选育对象的成活率, 这可能与连续多代采用活力较强的亲本进行繁殖有关, 甲壳动物的生长性能和成活率在选育过程中可能具有加性效应, 通过合理的选育策略可以使得甲壳动物的生长性能和成活率均得以提高(Campos-Montes et al, 2013; 郑静静, 2017)。在之前甲壳动物的选育工作中同样存在选育对象生长性能和成活率同时得到提升的现象, 例如三疣梭子蟹(Portunus trituberculatus)“黄选1号”子一代的体质量和成活率, 分别较未选育组提高2.91%和8.66% (李健等, 2013)。此外, 关于虾蟹类甲壳动物成活性状的遗传参数评估有较多报道, 已表明成活性状的遗传力一般属低遗传力(Gitterle et al, 2005; 栾生等, 2013; 高保全, 2014)。针对偏低遗传力性状选育时, 应将家系选择和个体选择结合起来实现更大的选择效应。
4 结论综上, 本研究结果表明: “长荡湖1号”选育第二代的两个品系在养殖性能方面已经获得了较好的遗传进展, 具有一定的推广应用潜力。“长荡湖1号”A系和B系在扣蟹阶段的最终平均体质量分别比对照组高11.07%和19.23%, 在成蟹阶段的最终平均体质量分别比对照组高14.04%和24.99%; 在成活率方面, A系在扣蟹和成蟹养殖阶段的最终成活率分别比对照组高6.77%和7.76%; A系和B系成蟹养殖的产量分别比对照组高23.87%和14.70%。
参考文献
王武, 王成辉, 马旭洲, 2013. 河蟹生态养殖[M]. 2版. 北京: 中国农业出版社, 59-84.
|
朱华平, 卢迈新, 黄樟翰, 等, 2010. 鱼类遗传改良研究综述[J]. 中国水产科学, 17(1): 168-181. |
庄振俊, 张冬冬, 姜晓东, 等, 2021. 中华绒螯蟹"长荡湖1号"子一代成蟹阶段养殖性能初步评估[J]. 南方水产科学, 17(3): 27-35. |
庄振俊, 张冬冬, 姜晓东, 等, 2022. 中华绒螯蟹"长荡湖1号"奇数年子一代的成蟹养殖性能评估[J]. 海洋渔业, 44(6): 747-758. |
农业农村部渔业渔政管理局, 水产技术推广站, 中国水产学会, 2021. 2021中国渔业统计年鉴[M]. 北京: 中国农业出版社, 24.
|
李健, 刘萍, 高保全, 等, 2013. 三疣梭子蟹新品种"黄选1号"的选育[J]. 渔业科学进展, 34(5): 51-57. |
杨帅帅, 吴仁福, 张保彦, 等, 2021. 大规格中华绒螯蟹亲本子代大眼幼体在微山湖地区的池塘扣蟹培育试验[J]. 水产科技情报, 48(2): 87-93. |
吴晓峰, 耿智, 冯广朋, 等, 2020. 长江口中华绒螯蟹成蟹资源现状及其繁殖特征[J]. 海洋渔业, 42(2): 205-213. |
邹杰, 马爱军, 王新安, 等, 2013. 鱼类育种技术研究进展[J]. 渔业信息与战略, 28(3): 199-207. DOI:10.13233/j.cnki.fishis.2013.03.008 |
张世勇, 傅洪拓, 乔慧, 等, 2013. 中华绒螯蟹遗传育种研究进展[J]. 中国农学通报, 29(20): 39-45. |
陈军伟, 马旭洲, 王武, 等, 2016. 不同规格中华绒螯蟹母本子代的生长特性比较[J]. 动物学杂志, 51(5): 895-906. |
范陈伟, 姜晓东, 吴旭干, 等, 2021. 不同规格中华绒螯蟹亲本子一代在扣蟹阶段养殖性能的比较研究[J]. 水产科技情报, 48(1): 33-39. |
林永淮, 龚忠智, 2009. 提高河蟹养殖成活率的四大关键措施[J]. 渔业致富指南, (8): 40-41. |
罗明, 周威, 王少兵, 等, 2020. "长荡湖1号"中华绒螯蟹选育子一代扣蟹养殖性能的初步研究[J]. 水产科技情报, 47(6): 324-328. |
郑静静, 2017. 日本囊对虾生长、存活及抗逆性状遗传参数的估计[D]. 湛江: 广东海洋大学: 17-20.
|
栾生, 罗坤, 阮晓红, 等, 2013. 凡纳滨对虾(Litopenaeus vannamei)体重、存活性状的遗传参数和基因型与环境互作效应[J]. 海洋与湖沼, 44(2): 445-452. |
高保全, 2014. 三疣梭子蟹(Portunus trituberculatus)遗传参数评估及新品种培育的研究[D]. 青岛: 中国海洋大学.
|
堵南山, 1998. 中华绒螯蟹的同属种类及其英文名称[J]. 水产科技情报, 25(3): 108-109, 113. |
董江水, 陈红军, 王新华, 等, 2007. 放养密度对河蟹育成规格、产量和成活率的影响[J]. 金陵科技学院学报, 23(4): 95-99. |
ARGUE B J, ARCE S M, LOTZ J M, et al, 2002. Selective breeding of Pacific white shrimp (Litopenaeus vannamei) for growth and resistance to Taura Syndrome Virus[J]. Aquaculture, 204(3/4): 447-460. |
BUSACK C A, GALL G A E, 1983. An initial description of the quantitative genetics of growth and reproduction in the mosquitofish, Gambusia affinis[J]. Aquaculture, 32(1/2): 123-140. |
CAMPOS-MONTES G R, MONTALDO H H, MARTÍNEZ-ORTEGA A, et al, 2013. Genetic parameters for growth and survival traits in Pacific white shrimp Penaeus (Litopenaeus) vannamei from a nucleus population undergoing a two-stage selection program[J]. Aquaculture International, 21(2): 299-310. |
EL-IBIARY H M, JOYCE J A, 1978. Heritability of body size traits, dressing weight and lipid content in channel catfish[J]. Journal of Animal Science, 47(1): 82-88. |
GITTERLE T, RYE M, SALTE R, et al, 2005. Genetic (co) variation in harvest body weight and survival in Penaeus (Litopenaeus) vannamei under standard commercial conditions[J]. Aquaculture, 243(1/2/3/4): 83-92. |
LONG X W, GUO Q, WANG X C, et al, 2020. Effects of fattening period on ovarian development and nutritional quality of adult female Chinese mitten crab Eriocheir sinensis[J]. Aquaculture, 519: 734748. |
LONG X W, WU X G, ZHU S C, et al, 2019. Salinity can change the lipid composition of adult Chinese mitten crab after long-term salinity adaptation[J]. PLoS One, 14(7): e0219260. |
PRESTON N P, CROCOS P J, KEYS S J, et al, 2004. Comparative growth of selected and non-selected Kuruma shrimp Penaeus (Marsupenaeus) japonicus in commercial farm ponds; implications for broodstock production[J]. Aquaculture, 231(1/2/3/4): 73-82. |
WANG S, HE Y, WANG Y Y, et al, 2016. Comparison of flavour qualities of three sourced Eriocheir sinensis[J]. Food Chemistry, 200: 24-31. |
WANG Q J, WU X G, LONG X W, et al, 2018a. Nutritional quality of different grades of adult male Chinese mitten crab, Eriocheir sinensis[J]. Journal of Food Science and Technology, 55(3): 944-955. |
WANG J, XU P, ZHOU G, et al, 2018b. Genetic improvement and breeding practices for Chinese mitten crab, Eriocheir sinensis[J]. Journal of the World Aquaculture Society, 49(2): 292-301. |
WU X G, ZHU S C, ZHANG H C, et al, 2020. Fattening culture improves the gonadal development and nutritional quality of male Chinese mitten crab Eriocheir sinensis[J]. Aquaculture, 518: 734865. |