 2024, Vol. 55
2024, Vol. 55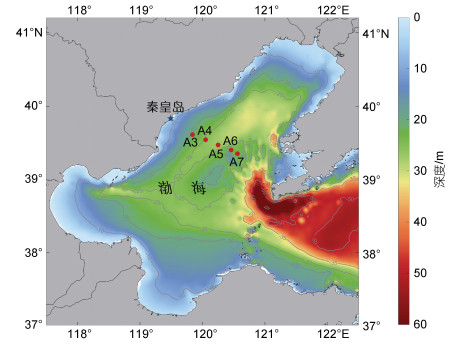


秦皇岛外海低氧区颗粒物吸收光谱特征

http://dx.doi.org/10.11693/hyhz20230700143
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 于廉涛, 张桂成, 杨伟, 赵亮, 李梦婷, 宋贵生. 2024.
- YU Lian-Tao, ZHANG Gui-Cheng, YANG Wei, ZHAO Liang, LI Meng-Ting, SONG Gui-Sheng. 2024.
- 秦皇岛外海低氧区颗粒物吸收光谱特征
- ABSORPTION SPECTRUM OF PARTICULATE MATTER IN LOW-OXYGEN AREA OFF THE QINHUANGDAO
- 海洋与湖沼, 55(1): 107-117
- Oceanologia et Limnologia Sinica, 55(1): 107-117.
- http://dx.doi.org/10.11693/hyhz20230700143
文章历史
-
收稿日期:2023-07-10
收修改稿日期:2023-09-26
引用本文
于廉涛, 张桂成, 杨伟, 赵亮, 李梦婷, 宋贵生. 2024. 秦皇岛外海低氧区颗粒物吸收光谱特征[J]. 海洋与湖沼, 55(1): 107-117

YU Lian-Tao, ZHANG Gui-Cheng, YANG Wei, ZHAO Liang, LI Meng-Ting, SONG Gui-Sheng. 2024. ABSORPTION SPECTRUM OF PARTICULATE MATTER IN LOW-OXYGEN AREA OFF THE QINHUANGDAO[J]. Oceanologia et Limnologia Sinica, 55(1): 107-117.
秦皇岛外海低氧区颗粒物吸收光谱特征
1. 天津大学海洋科学与技术学院 天津市海洋环境与安全保障重点实验室 天津 300072;
2. 天津科技大学海洋与环境学院 天津 300457
2. 天津科技大学海洋与环境学院 天津 300457
摘要:近年来, 渤海夏季低氧现象频发, 引起了人们的广泛关注。然而对该海域低氧形成的机制还未得到充分认识。研究基于在秦皇岛外海的现场观测, 分析了海水中颗粒物吸收光谱特征及其与不同粒径浮游植物叶绿素a (chl a)组成、环境因子的关系, 评估了夏季底层水体脱氧过程中有机物来源与特征。结果显示, 夏季秦皇岛外海微型浮游植物chl a占总量的80%。表层水体中, 总颗粒物吸收光谱[ap(λ)]特征由浮游植物色素吸收光谱[aph(λ)]主导, 在中、底层水体中则由碎屑颗粒物吸收光谱[ad(λ)]主导。垂向上, ap(440)和ad(440)均表现为表层<中层<底层。结果还表明, 浮游植物粒径主导秦皇岛外海chl a的光吸收效率, 即aph*(440)。基于三粒级chl a含量, 可利用多元回归预测aph(440)。碎屑颗粒物的吸收光谱同样受浮游植物群落、有机质相对含量等的影响。研究结果表明初级生产产生的微型颗粒有机物是底层水体脱氧的主要底物。
关键词:颗粒物 吸收光谱 叶绿素a 秦皇岛外海
ABSORPTION SPECTRUM OF PARTICULATE MATTER IN LOW-OXYGEN AREA OFF THE QINHUANGDAO
1. Tianjin Key Laboratory for Marine Environmental Research and Service, School of Marine Science and Technology, Tianjin University, Tianjin 300072, China;
2. College of Marine and Environmental Sciences, Tianjin University of Science and Technology, Tianjin 300457, China
2. College of Marine and Environmental Sciences, Tianjin University of Science and Technology, Tianjin 300457, China
Abstract: More and more attention has been paid on the oxygen depletion in the Bohai Sea in recent years. However, the mechanism of deoxygenation has not been well known. We investigated the characterization of the absorption spectra of suspended particles in seawater off Qinhuangdao, Hebei, China. Furthermore, and evaluated the sources and features of organic matter responsible for deoxygenation in bottom water in summer based on the relationships of particulate absorption spectra with the size-based components of chlorophyll a (chl a) and with environment factors. Results show that nano-phytoplankton contributed approximately 80% of total chl a. The absorption spectra of pigment [aph(λ)] reflected the major feature of total particulate absorption spectra [ap(λ)]in surface water, while that of detritus [ad(λ)] was dominant in middle and bottom layers. ap(440) and ad(440) increased from surfac to bottom waters. The size of phytoplankton controlled the absorption efficiency of pigment, i.e., aph*(440). Therefore, aph(440) could be calculated using multiple regression based on Chl a concentrations of three phytoplankton size classes. The characters of ad(λ) were also influenced by the phytoplankton community and the relative abundance of organic detritus. This study revealed that nano-sized organic particles produced during primary production were the main substrates of deoxygenation in summer in Bohai Sea.
Key words:
particles absorption spectra chlorophyll a offshore of Qinhuangdao
悬浮颗粒物(suspended particulate matter, SPM)是海水的重要组成部分, 也是海洋物质循环的重要载体(Hong et al, 1999; Che et al, 2003), 在调控海洋生物地球化学循环、生态过程以及碳埋藏过程中均起到重要作用(Reynolds et al, 2021)。同时, 海水中SPM是水柱、沉积物和食物链之间重要的物质交换纽带(Turner et al, 2002), 对海水中物质的迁移转化有重要意义。SPM可分为有机组分和无机组分: 无机组分主要包括陆源输入的各类无机矿物碎屑、生物过程产生的硅质、钙质骨架碎屑等以及海水化学过程中产生的次生矿物; 有机组分主要包括浮游植物、浮游动物、细菌、生物残骸及其代谢产物(Goldberg, 1979)。
SPM通过吸收和反射影响太阳辐射在海水中的穿透深度, 其吸收光谱[ap(λ)]可分为浮游植物吸收光谱[aph(λ)]和碎屑颗粒物吸收光谱[ad(λ)]。aph(λ)在400~450 nm和675 nm处分别存在显著的色素吸收峰(Bricaud et al, 1990)。aph(λ)主要受叶绿素a (chl a)浓度影响, 二者存在一定的正相关关系(Sathyendranath et al, 1988; Bricaud et al, 1995)。然而, 其吸收光谱的形状还取决于藻类细胞中光合色素的吸收特性以及一些其他因素, 如浮游植物种群结构、生长周期、光照以及包裹效应等(Bricaud et al, 1995, 1998; Stramski et al, 2002)。另外, 浮游植物吸收光谱存在一定的时空变化(Babin et al, 2003)。其中, 包裹效应与藻类细胞粒径和色素浓度有关, 色素浓度越高, 细胞粒径越大, 包裹效应越强(Morel et al, 1981; Bricaud et al, 1990)。因此可将aph(λ)对chl a浓度进行归一化, 得到浮游植物比吸收aph*(λ)(Bricaud et al, 1995), 用以描述水体中浮游植物的吸收特性, 判别浮游植物大小及组成(周雯等, 2012; Sun et al, 2022)。研究发现辅助色素在aph*(440)变化中扮演重要角色, 因此常采用440 nm波长下的比吸收作为参考。
在开阔大洋, 碎屑颗粒物主要源自浮游植物产生的有机碎屑, 因此, ad(λ)往往小于aph(λ); 在河口和陆架海中, 陆源输入有机颗粒物和无机矿物的影响导致ad(λ)波动较大(Woźniak et al, 1992)。ad(λ)与SPM浓度之间存在正相关关系(Bowers et al, 1996; Babin et al, 2003), 随波长呈指数衰减特征(Bricaud et al, 1998):
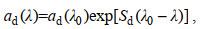 (1)
(1)其中λ0是参考波长, Sd是光谱斜率, 反映ad随波长降低的程度。Sd与无机矿物和有机质的比例有关, 变化幅度较小(Babin et al, 2003), 且会受到有机碎屑组成的影响(Bricaud et al, 2010)。此外, 邢小罡等(2008)在渤海海域的研究发现ad与Sd存在明显的负相关关系。
水体层化和底层水体中有机物大量分解耗氧是陆架海低氧形成的基本条件(Rabalais et al, 2007), 而有机物分解耗氧又分为水柱耗氧和沉积物耗氧两部分(Dortch et al, 1994; Song et al, 2020)。水柱耗氧的贡献主要受控于水体中颗粒有机物的逗留时间。如在密西西比河口、长江口等浅水区, 通常由沉积物耗氧主导(Morse et al, 2007; Hetland et al, 2008; Zhang et al, 2017), 在圣劳伦斯河口等深水区(Bourgault et al, 2012), 由于颗粒有机物(particulate organic matter, POM)在水柱中停留时间较长, 以水柱耗氧为主; 然而, 在切萨皮克湾(Li et al, 2015)和路易斯安娜湾(Corbett et al, 2004; Green et al, 2006; Hetland et al, 2008)等浅水区, 由于水平流通量较大, 阻碍了POM的沉降, 导致其以水柱耗氧为主。
渤海是我国典型的半封闭型浅海, 三面环陆, 水体交换能力较弱(魏皓等, 2002)。受人为活动的显著影响, 渤海SPM浓度较高(Qiu et al, 2016)。近年来, 渤海洼地底层水体在夏季存在明显的酸化和低氧(Zhai et al, 2012; 张华等, 2016; Zhao et al, 2017; 李志成等, 2021)。Song等(2020)研究发现秦皇岛外海水柱耗氧主导了该区域底层水低氧的形成, 这与以往对浅水区以沉积物耗氧为主的认识不同(Morse et al, 2007; Hetland et al, 2008; Zhang et al, 2017)。在全球气候变化背景下, 渤海甲、硅藻比逐渐升高, 且浮游植物粒径向微型和微微型过渡(杨阳等, 2016; 赵相伟等, 2020), 因此, 浮游植物小型化导致POM沉降速率减小是引起渤海水柱耗氧主导的主要原因(Wei et al, 2021)。然而, 目前对该区域有机物来源、组成和特性的认识还不足。因此, 本文研究了夏季秦皇岛外海海域颗粒物吸收光谱的时空变化特征, 量化了aph(λ)与chl a浓度和组成的关系, 探讨了影响颗粒物吸收光谱的主要环境因子。本研究结果有助于进一步认识颗粒物来源和组成对低氧形成的潜在影响。
1 样品采集与测定 1.1 样品采集海水样品分别于2021年6月20日、7月20日、8月30日沿秦皇岛海域一个近岸-外海断面采集(图 1, 表 1)。每个站位分别采集表层、中层和底层海水样品, 其中, 表层水取3 m水深; 中层依据站位水深设定, 一般为8~12 m之间; 底层以离底约2 m深度处采集。海水样品由Seabird CTD rosette(SBE19 plus)系统携带的4 L Niskin采样器采集后, 立即转移至4 L洁净塑料桶中, 避光保存, 并尽快过滤。海水温、盐和溶解氧(DO)剖面由温盐深剖面仪(CTD)测定。
表 1 航次具体信息 Tab. 1 Latitude, longitude and water depth of sampling stations
| 站位 | 纬度/°N | 经度/°E | 水深/m |
| A3 | 39.617 | 119.833 | 21 |
| A4 | 39.533 | 120.033 | 25 |
| A5 | 39.467 | 120.250 | 26 |
| A6 | 39.400 | 120.450 | 22 |
| A7 | 39.367 | 120.533 | 22 |
一部分海水样品经焙烧过(450 ℃, 4 h)的25 mm GF/F (Whatman)滤膜过滤并记录过滤体积。滤膜置于塑料培养皿内, 并采用铝箔纸包裹, 用于测定颗粒物吸收光谱。滤液转移至250 mL聚丙烯(PP)样品瓶, 用于测定营养盐浓度。剩余样品分别经孔径为20 μm筛绢、2 μm尼龙滤膜(Pall Life Science)和GF/F(Whatman)滤膜过滤一定体积后, 滤膜采用铝箔纸包裹, 用于分析不同粒级chl a浓度。所有颗粒物和溶解态样品采集后, 立即置于−20 ℃冰箱中冷冻保存。
1.2 样品分析 1.2.1 颗粒物吸收光谱颗粒物吸收光谱采用紫外可见分光光度计(PerkinElmer, LAMBDA850)测定。仪器搭配150 mm积分球, 扫描范围750~300 nm, 步长1 nm。样品测定前, 为防止细胞破裂, 滴加1~2滴经0.2 µm孔径滤膜过滤的对应站位海水, 在室温下避光解冻、升至室温。每个样品测定一次吸收光谱后, 将滤膜旋转120°后继续测定, 共平行测定三次, 对应波长的平均吸光度值记为样品的吸光度[ODsample(λ)]。以同批次, 未过滤样品的GF/F滤膜滴加1~2滴MilliQ水, 测定其吸光度为滤膜空白[ODblank(λ)]。
三次测定后, 滤膜经10 mL甲醇(Merck)于暗处萃取18~24 h, 以除去浮游植物色素。萃取后, 再次过滤并采用MilliQ水冲洗去除甲醇, 以上述相同步骤三次测定的平均结果为碎屑颗粒物吸光度。总颗粒物和碎屑吸收光谱依据下式计算(Röttgers et al, 2012):
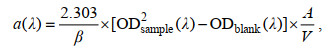 (2)
(2)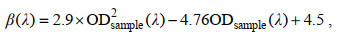 (3)
(3)其中a(λ)为总颗粒物或碎屑在波长λ处吸收系数(单位: m–1), A为滤膜有效面积(单位: m2), V为过滤样品体积(单位: m3), β(λ)为波长λ处的放大系数(无量纲)。
浮游植物的吸收系数由二者相减得到:
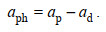 (4)
(4)Chl a样品经90%的丙酮溶液萃取24 h后采用荧光计(Trilogy)测定(Parsons et al, 1984), 20 μm筛绢、2 μm尼龙和GF/F滤膜截流样品分别代表小型(Micro, > 20 μm)、微型(Nano, 2~20 μm)、微微型(Pico, 0.7~2 μm)浮游植物的chl a。营养盐依据Hansen(1999)的方法, 使用AA3营养盐自动分析仪(Seal, 英国)测定。
2 结果 2.1 水文、生物和化学特征夏季, 秦皇岛外海水体呈现出明显的分层现象, 水柱分为上、下混合层和中层跃层(图 2), 与以往观测一致(Song et al, 2020)。6、7月跃层较浅, 8月加深, 且8月份在距离中央浅滩较近区域(A6和A7站位)无明显分层现象(图 2c, 2f, 2i), 反映水体层化将逐渐破坏。在整个夏季海水温度逐月升高, 盐度逐月下降, 6~8月上混合层平均温度从(20.79±0.54) ℃上升至(25.54±0.20) ℃, 盐度从31.74±0.05降至30.78±0.20; 下混合层平均温度从(13.16±1.67) ℃上升至(21.37± 2.10) ℃, 盐度从32.06±0.21下降至31.16±0.30。温盐均在上下混合层均呈现显著的差异, 7月份差异最为显著, 表底温度差范围在7.35~12.59 ℃, 盐度差范围在0.34~1.22。
 |
| 图 2 2021年夏季温度(a~c)、盐度(d~f)、DO (g~i)断面分布 Fig. 2 Temperature (a~c), salinity (d~f), and DO (g~i) cross-sectional distribution in summer 2021 注: 图中黑色实线代表跃层上下边界, 跃层位置通过计算浮力频率的平方(N2≥10−3)确定(Gill, 1982) |
整个夏季chl a在中层最高, 且月际间无显著变化[(3.48±0.58)~(3.92±2.52) µg/L]。表层从(1.76±0.56) µg/L上升到(3.16±1.23) µg/L; 底层趋势与表层相反, 从(3.94±0.60) µg/L下降至(1.79±0.99) µg/L。研究期间微型浮游植物主导chl a, 占总chl a浓度的78.3%± 14.1%, 小型和微微型比例相当, 分别为10.1%±10.5%和11.7%±8.9% (图 3)。微微型浮游植物在研究期间并无明显的时间变化, 但存在较为显著的空间变化, 小型浮游植物在8月占比显著增加(图 3), 在A3站位达到约40%, 与微型浮游植物贡献相当。垂向上小型浮游植物占比在底层显著升高, 其中7月最高, 平均为表层的4倍; 8月份最低, 平均为表层1.7倍。微型、微微型无明显垂向差异。由于光合作用, 表层和中层海水中DO始终处于饱和或过饱和状态。底层海水中DO平均浓度自6月[(7.6±0.2) mg/L]开始逐渐降低[8月: (5.0±0.2) mg/L](图 2g, 2h, 2i), 其饱和度则由88.9%±2.0%降低至66.5%±7.7%。8月底DO浓度和饱和度均显著高于以往年份(Zhai et al, 2012; Song et al, 2020), 表明该年份水体净耗氧速率较低。
 |
| 图 3 各月份表、中、底层分级叶绿素占比 Fig. 3 The proportion of chl a in the surface, middle, and bottom waters |
图 4给出了三个月份表、中、底层总颗粒物、浮游植物和碎屑颗粒物的吸收光谱。aph(λ)在435~ 445 nm具有一个吸收肩, 且在675 nm呈现chl a特征吸收峰, 与以往研究结果一致(Bricaud et al, 1990;Tassan et al, 1995; 刘洋洋等, 2015)。ad(λ)随波长增加而逐渐降低。ap(λ)在不同水层存在显著的差异, 在表层与aph(λ)相似, 而在底层与ad(λ)相近, 表明上层水体中, 活体浮游植物在总颗粒物中占主导, 而中、底层水体中则是碎屑颗粒物为主。aph*(λ)光谱特征(图 5)与aph(λ)(图 4)类似, 但在8月A3站位表、中层(图 5a, 5b红色点划线) aph*(λ)与其他站位呈现显著差异, 在蓝光波段吸收峰左移, 且肩峰消失, 呈现较为平滑的曲线, 整体与Sun等(2022)研究中硅藻的aph*(λ)吸收特征较为相似。
 |
| 图 4 秦皇岛外海表层(a, d, g)、中层(b, e, h)和底层(c, f, i)海水中总颗粒物、碎屑与浮游植物吸收光谱 Fig. 4 Absorption spectras of SPM, nonalgal particles and phytoplankton for surface (a, d, g), middle (b, e, h), and bottom (c, f, i) waters off Qinhuangdao |
 |
| 图 5 秦皇岛外海表层(a)、中层(b)和底层(c)浮游植物比吸收光谱 Fig. 5 Specific absorption coefficients of phytoplankton in surface (a), middle (b), and bottom (c) waters off Qinhuangdao 注: 图中红色线为8月A3站位 |
夏季, ap(440)和ad(440)均表现为表层<中层<底层(图 5a, 5b)。在7、8月, aph(440)在底层水体显著低于表层和中层, 底层水体aph(440)仅占其上层平均值的25% (图 5c)。总体而言, ap(440)在表层和逐月增加, 中层逐月减少, 底层无显著变化[平均值: (0.40±0.07) m–1], 浮游植物群落结构变化导致的沉降速率改变和跃层加深伴随的颗粒物累积深度的下移可能是导致表中层水体中ap(440)月变化的主要因素。此外, 底层水体中ad(440)的增加表明碎屑颗粒物在底层水体中存在累积, 为该海区夏季底层水体脱氧(Zhai et al, 2012; Song et al, 2020)提供了底物。aph*(440)在7月表、中层显著高于6、8月, 底层呈现相反情况, 而6、8月间未见明显的变化趋势, 表、中层维持在0.030 m2/mg上下, 底层维持在0.020 m2/mg上下, 7月份 aph*(440)与剩余月份的差异表明浮游植物群落构成出现变动。多数研究结果表明, 浮游植物比吸收系数的分布具有明显的区域特征, 在本次观测期间aph*(440)变化范围在0.010~0.060 m2/mg, 平均为0.032 m2/mg, 远低于南海北部海区测量结果(0.041~ 0.295 m2/mg, 均值0.137 m2/mg)(王桂芬等, 2005); 与加利福尼亚湾南部海区(0.008~0.102 m2/mg, 均值0.041 m2/mg)(Millán-Núñez et al, 2004)和长江口临近海域(春季0.016~0.123 m2/mg, 均值0.040 m2/mg; 夏季0.012~0.064 m2/mg, 均值0.029 m2/mg)(刘洋洋, 2015)测量结果接近。
 |
| 图 6 ap(440) (a)、ad(440) (b)、aph(440) (c)和aph*(440) (d)在表、中和底层海水中的平均含量 Fig. 6 Average contents of ap(440) (a), ad(440) (b), aph(440) (c) and aph*(440) (d) in surface, middle and bottom layers 注: 误差棒代表相对标准偏差 |
总体上, aph(440)随chl a升高而升高(P<0.01)(图 7a)。在表、中层水体中, aph(440)与chl a在6月和7月分别呈现显著相关性, 且当chl a浓度相同时, 7月整体呈现更高的aph(440)(图 7b), 表明7月份浮游植物色素对光的吸收效率更高。以往研究表明, 颗粒物包裹效应与色素构成均能够显著影响aph(440) (Sathyendranath et al, 1987; Greg Mitchell et al, 1988; Bricaud et al, 1990; Babin et al, 1993)。其中, 包裹效应会降低浮游植物光吸收能力(Stuart et al, 1998; Bricaud et al, 2004), 即活体浮游植物粒径越小, 单位chl a的光吸收越强。表、中层上、中层水体中, 7月aph*(440)显著高于6、8月, 分别达到(0.050±0.007)和(0.041±0.012) m2/mg (图 7), 且7月小型浮游植物占比(2.5%±2.0%)显著低于6月(6.0%±3.0%)和8月(15.1%±12.5%)(图 3)。此外, 8月份各站位间浮游植物构成差异显著(图 3), 可能是导致aph(440)与chl a无明显关系(图 7b)的主要原因。夏季, 渤海浮游植物为甲藻和硅藻共同占优, 而秋季则以硅藻为绝对优势种(杨阳等, 2016)。8月小型浮游植物占比的大幅提升(图 3), 反映出夏末优势藻种从甲、硅藻共同占优逐步向硅藻为优势种的转变过程。综合整个夏季表、中层样品, 去除浮游植物构成与其他样品差异较大的样品, 划分为数量相近的两组。发现在小型浮游植物(Micro)占总叶绿素含量≤5%和5%~15%时, aph(440)与chl a分别呈显著的线性相关, aph(440)在小型浮游植物叶绿素占比更小时(≤5%)的平均值比高占比(5%~15%)时高6% (图 7c), 进一步佐证浮游植物粒径是影响该海区夏季浮游植物光吸收效率的主要因素。
 |
| 图 7 所有样品(a), 表、中层样品(b)及小型浮游植物占比小于15%样品(c)aph(440)与chl a关系图 Fig. 7 Affection plots between aph(440) and chl a of all samples (a), surface and middle samples (b) and Micro phytoplankton less than 15% samples 注: b图中蓝色和橙色直线分别为6月和7月线性拟合结果[6月: aph(440)=0.033chl a−0.004, R2=0.90, P<0.01; 7月: aph(440) = 0.045chl a+ 0.005, R2=0.75, P<0.01], 阴影代表 95%置信区间, c图中FMicro表示小型浮游植物占比, 蓝色和红色直线分别为小型浮游植物占总chl a 5%~15%和≤5%时的拟合结果[5%<FMicro<15%: aph(440)=0.031chl a+0.008, R2=0.61, P<0.01; FMicro≤5%: aph(440)=0.032chl a+0.023, R2=0.60, P<0.01] |
考虑到不同粒径浮游植物对光吸收的影响, 仅靠单一粒径藻类无法对aph(440)进行量化, 因此, 本研究将三种浮游植物(Pico、Nano、Micro)叶绿素a浓度与aph(440)之间相关关系, 依据公式(6)进行多元回归模型求得三种浮游植物比吸收系数:
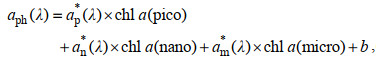 (6)
(6)式中ap*、an*、am*为三种浮游植物的比吸收系数, chl a (pico)、chl a(nano)、chl a(micro)为三种浮游植物叶绿素a浓度, 440 nm波长拟合结果分别为0.033、0.031、0.026 m2/mg。b为常数项, 结果为−0.003。对比刘洋洋(2015)对长江口颗粒物的三粒级模型模拟结果[ap*(440)=0.088 m2/mg、an*(440)=0.017 m2/mg、am*(440)=0.022 m2/mg], 本研究显著低估了ap*(440), 但拟合结果更符合浮游植物粒径越大, 光吸收效率越低的规律(Stuart et al, 1998; Bricaud et al, 2004)。造成模型结果之间差异的原因可能是浮游植物群落构成上的差异, 由于pico浮游植物在整个观测期间占比较少而导致对其一定程度上的低估。模型得到的aph(440)在0~0.25 m–1范围内, 拟合结果比实测值整体偏低(图 8), 标准偏差和相对误差分别为(0.023± 0.022) m–1和22%, 略大于测量标准偏差(0.015±0.021) m–1和相对误差(< 14%)(Röttgers et al, 2012, 本文)。除对ap*(440)的低估外, 本研究用于多元回归的数据量少(28个), 且只考虑了浮游植物粒径大小, 忽略了色素组成对aph(440)的影响(Hoepffner et al, 1992; Woźniak et al, 1992; Sathyendranath et al, 2001), 是导致拟合结果偏低的原因。
 |
| 图 8 多元回归模型aph(440)结果与实测值比较 Fig. 8 Comparison of multiple regression model aph(440) results with the measured values |
6至7月, 研究海域水体中ad(440)与无机营养盐无明显相关性(P > 0.05), 然而, ad(440)与硅酸盐(DSi)在8月呈显著的正相关关系(图 9)。该结果表明, 8月升高的小型浮游植物以硅藻为主, 且硅藻在光合作用过程中利用海水中DSi, 浮游植物死后其尸体在沉降过程中, 生物硅逐渐溶解提高水体中DSi浓度。因此, 8月底层水体富含生物硅的碎屑颗粒物(图 6b)是硅酸盐的主要底物。以A3站为例, 小型浮游植物叶绿素占总量的37%以上, 是该航次其他站位平均水平的2.5~5.7倍。A3站靠近秦皇岛扇贝养殖区, 贝类摄食能够促使浮游植物群落结构向生长速率较快的硅藻转变(Olsson et al, 1992), 且秦皇岛近海养殖扇贝对甲藻的摄食选择性显著高于硅藻(丁任业等, 2023)。以上结果表明在A3站, 大量硅藻的生长利用了水体中的DSi, 导致表、中层水体中DSi浓度显著降低(0.45和0.19 µmol/L), 且存在明显的硅限制(Si: N<0.7, Si: P<7)(Justić et al, 1995)。
 |
| 图 9 8月ad (440)与DSi相关性 Fig. 9 Correlation between ad (440) and DSi in August 注: 阴影部分代表 95%置信区间, aph(440)=0.014(chl a)2−0.023chl a +0.049, R2=0.95, P<0.01 |
Sd在整个研究期间保持相对稳定, 变化范围为0.008 4~0.011 7 nm–1, 整体略低于2008年邢小罡等(2008)测量结果。由于Sd与无机矿物和有机质的比例有关, 有机质占比更高的颗粒物通常具有更高的Sd (Babin et al, 2003), 各水层中, 6~7月Sd高于8月(表 2), 表明该期间内水体中有机质相对含量最高。该结果进一步表明6~7月上层水体高的初级生产向底层水体中输送了大量颗粒有机碎屑, 且在水柱中不断累积(见3.1节), 为夏季该海区显著的水柱耗氧(Song et al, 2020)提供了底物。至8月, Sd略有降低, 这与浮游植物群落向硅藻转变有关, 即硅藻体中生物硅含量较高, 降低了碎屑中有机质含量。
表 2 秦皇岛外海6、7、8月各水层碎屑光谱斜率 Tab. 2 Spectral slopes of nonalgal particles in each water layer of study area in June, July and August
| 月份 | Sd /nm–1 | ||
| 表层 | 中层 | 底层 | |
| 6月 | 0.010 0±0.000 2 | 0.010 0±0.000 5 | 0.010 1±0.000 2 |
| 7月 | 0.010 4±0.000 5 | 0.010 2±0.000 6 | 0.010 3±0.000 0 |
| 8月 | 0.009 6±0.000 8 | 0.010 0±0.001 0 | 0.009 7±0.000 5 |
为进一步探究颗粒物对低氧形成的影响, 我们对数据集进行主成分分析(PCA), 包括吸收系数、营养盐、温盐、叶绿素、DO共11项指标(图 10)。PCA中各变量均按比例缩放到−1~1, 结果表明前两个主成分解释了76%的变异性。在PC1下, DO及其饱和度(DO%)与ap(440)、ad(440)间呈现强烈的负相关性, 表明初级生产过程中产生的有机碎屑在底层水体中分解, 是DO消耗的主控因子, 与Song等(2020)估算结果一致; DO%与aph(440)呈现强烈的正相关关系, 表明浮游植物光合作用对DO%的贡献。在PC2下, DO及其DO%与营养盐呈现强负相关, 这与富营养化导致浮游植物快速繁殖, 其死亡沉降产生大量有机物分解, 从而消耗DO形成低氧的认识一致(Rabalais et al, 2010; McCarthy et al, 2013; Zhang et al, 2017)。
 |
| 图 10 基于吸收系数、营养盐、温盐、叶绿素、DO等11项指标主成分分析 Fig. 10 The principal component analysis (PCA) based on 11 indexes, include absorption coefficient, nutrient, temperature, salinity, chlorophyll and DO 注: 图中SiO44–、NOx–、PO43–分别代表硅酸盐浓度、硝酸及亚硝酸盐浓度和磷酸盐浓度 |
基于现场观测结果表明, 夏季秦皇岛外海水体呈现明显的层化。Chl a以微型浮游植物为主, 占chl a总量的80%。8月下旬, 尤其是近岸海域小型浮游植物(硅藻)占比升高。aph(λ)呈现明显的色素吸收峰, 而ad(λ)随波长呈指数递减, ap(λ)在表层水体中的特征与aph(λ)相似, 在中层和底层水体中与ad(λ)相近, 表明表层颗粒物以活体浮游植物为主, 在中层和底层则以碎屑颗粒物为主。ad(440)随深度逐渐升高, 该结果主导了ap(440)的垂直分布特征: 表层<中层<底层。在底层, ad(440)逐月升高, 表明碎屑颗粒物在水柱中不断积累, 为底层水体耗氧提供了丰富的底物。aph(440)与chl a的线性相关性及三粒级模型的拟合结果表明浮游植物粒径主导了秦皇岛外海chl a的光吸收效率, 同时浮游植物色素组成会调控二者的相关性。碎屑颗粒物的吸收光谱同样受浮游植物群落、有机质相对含量等因素的影响。研究结果还表明夏季该海区初级生产产生的新鲜颗粒有机物是水体脱氧的主要底物。
参考文献
丁任业, 查道军, 谢艺萱, 等, 2023. 秦皇岛贝类养殖区4种贝类对浮游植物的摄食选择性[J]. 烟台大学学报(自然科学与工程版), 36(1): 66-72. |
王桂芬, 曹文熙, 许大志, 等, 2005. 南海北部水体浮游植物比吸收系数的变化[J]. 热带海洋学报, 24(5): 1-10. DOI:10.3969/j.issn.1009-5470.2005.05.001 |
邢小罡, 赵冬至, 刘玉光, 等, 2008. 渤海非色素颗粒物和黄色物质的吸收特性研究[J]. 海洋环境科学, 27(6): 595-598. |
刘洋洋, 2015. 长江口邻近海域赤潮水体光吸收特性研究[D]. 上海: 华东师范大学.
|
刘洋洋, 沈芳, 李秀珍, 2015. 长江口邻近海域赤潮水体浮游植物光吸收特性分析[J]. 环境科学, 36(6): 2019-2027. DOI:10.13227/j.hjkx.2015.06.015 |
李志成, 魏皓, 张海彦, 等, 2021. 渤海夏季底层氧亏损分布的年际差异分析[J]. 海洋与湖沼, 52(3): 601-613. |
杨阳, 孙军, 关翔宇, 等, 2016. 渤海网采浮游植物群集的季节变化[J]. 海洋通报, 35(2): 121-131. |
张华, 李艳芳, 唐诚, 等, 2016. 渤海底层低氧区的空间特征与形成机制[J]. 科学通报, 61(14): 1612-1620. |
周雯, 孙兆华, 曹文熙, 等, 2012. 浮游植物的吸收-衰减特性及其与粒径间的关系[J]. 光谱学与光谱分析, 32(12): 3347-3352. DOI:10.3964/j.issn.1000-0593(2012)12-3347-06 |
赵相伟, 魏玉秋, 孙军, 等, 2020. 春、夏季秦皇岛海域超微型浮游植物[J]. 海洋学报, 42(2): 106-114. |
魏皓, 田恬, 周锋, 等, 2002. 渤海水交换的数值研究-水质模型对半交换时间的模拟[J]. 青岛海洋大学学报, 32(4): 519-525. |
BABIN M, STRAMSKI D, FERRARI G M, et al, 2003. Variations in the light absorption coefficients of phytoplankton, nonalgal particles, and dissolved organic matter in coastal waters around Europe[J]. Journal of Geophysical Research: Oceans, 108(C7): 3211. |
BABIN M, THERRIAULT J C, LEGENDRE L, et al, 1993. Variations in the specific absorption coefficient for natural phytoplankton assemblages: impact on estimates of primary production[J]. Limnology and Oceanography, 38(1): 154-177. DOI:10.4319/lo.1993.38.1.0154 |
BOURGAULT D, CYR F, GALBRAITH P S, et al, 2012. Relative importance of pelagic and sediment respiration in causing hypoxia in a deep estuary[J]. Journal of Geophysical Research: Oceans, 117(C8): C08033. |
BOWERS D G, HARKER G E L, STEPHAN B, 1996. Absorption spectra of inorganic particles in the Irish Sea and their relevance to remote sensing of chlorophyll[J]. International Journal of Remote Sensing, 17(12): 2449-2460. DOI:10.1080/01431169608948782 |
BRICAUD A, BABIN M, CLAUSTRE H, et al, 2010. Light absorption properties and absorption budget of Southeast Pacific waters[J]. Journal of Geophysical Research: Oceans, 115(C8): C08009. |
BRICAUD A, BABIN M, MOREL A, et al, 1995. Variability in the chlorophyll-specific absorption coefficients of natural phytoplankton: analysis and parameterization[J]. Journal of Geophysical Research: Oceans, 100(C7): 13321-13332. DOI:10.1029/95JC00463 |
BRICAUD A, CLAUSTRE H, RAS J, et al, 2004. Natural variability of phytoplanktonic absorption in oceanic waters: influence of the size structure of algal populations[J]. Journal of Geophysical Research: Oceans, 109(C11): C11010. |
BRICAUD A, MOREL A, BABIN M, et al, 1998. Variations of light absorption by suspended particles with chlorophyll a concentration in oceanic (case 1) waters: analysis and implications for bio-optical models[J]. Journal of Geophysical Research: Oceans, 103(C13): 31033-31044. DOI:10.1029/98JC02712 |
BRICAUD A, STRAMSKI D, 1990. Spectral absorption coefficients of living phytoplankton and nonalgal biogenous matter: a comparison between the Peru upwelling areaand the Sargasso Sea[J]. Limnology and Oceanography, 35(3): 562-582. DOI:10.4319/lo.1990.35.3.0562 |
CHE Y, HE Q, LIN W Q, 2003. The distributions of particulate heavy metals and its indication to the transfer of sediments in the Changjiang Estuary and Hangzhou Bay, China[J]. Marine Pollution Bulletin, 46(1): 123-131. DOI:10.1016/S0025-326X(02)00355-7 |
CORBETT D R, MCKEE B, DUNCAN D, 2004. An evaluation of mobile mud dynamics in the Mississippi River deltaic region[J]. Marine Geology, 209(1/2/3/4): 91-112. |
DORTCH Q, RABALAIS N N, TURNER R E, et al, 1994. Respiration rates and hypoxia on the Louisiana shelf[J]. Estuaries, 17(4): 862-872. DOI:10.2307/1352754 |
GILL A E, 1982. Atmosphere-Ocean Dynamics[M]. London: Academic Press, 117-188.
|
GOLDBERG E D, 1979. Riley, J. P., and R. Chester [Eds. ]. 1978. Chemical oceanography, v. 7, 2nd Edition. Academic Press, London, xvi + 508 p. £19.60; US$40.50. [J]. Limnology and Oceanography, 24(6): 1178-1178.
|
GREEN R E, BIANCHI T S, DAGG M J, et al, 2006. An organic carbon budget for the Mississippi River turbidity plume and plume contributions to air-sea CO2 fluxes and bottom water hypoxia[J]. Estuaries and Coasts, 29(4): 579-597. DOI:10.1007/BF02784284 |
GREG MITCHELL B, KIEPER D A, 1988. Variability in pigment particulate fluorescence and absorption spectra in the northeastern Pacific Ocean. Deep Sea Research Part A[J]. Oceanographic Research Papers, 35(5): 665-689. |
HANSEN H P, KOROLEFF F, 1999. Determination of nutrients [M]//GRASSHOFF K, KREMLING K, EHRHARDT M. Methods of Seawater Analysis. 3rd ed. New York: Wiley-VCH: 159-228.
|
HETLAND R D, DIMARCO S F, 2008. How does the character of oxygen demand control the structure of hypoxia on the Texas–Louisiana continental shelf?[J]. Journal of Marine Systems, 70(1/2): 49-62. |
HOEPFFNER N, ALBRECHT H P, HAAGEN G, et al, 1992. Sonderform einer akrokeratose bazex bei kleinzelligem bronchialkarzinom[J]. Hautarzt, 43: 496-499. |
HONG H S, CHEN W Q, XU L, et al, 1999. Distribution and fate of organochlorine pollutants in the pearl river estuary [J]. Marine Pollution Bulletin, 39(1/2/3/4/5/6/7/8/9/10/11/12): 376-382.
|
JUSTIĆ D, RABALAIS N N, TURNER R E, et al, 1995. Changes in nutrient structure of river-dominated coastal waters: stoichiometric nutrient balance and its consequences [J]. Estuarine, Coastal and Shelf Science, 40(3): 339-356.
|
LI Y, LI M, KEMP W M, 2015. A Budget analysis of bottom-water dissolved oxygen in Chesapeake Bay[J]. Estuaries and Coasts, 38(6): 2132-2148. DOI:10.1007/s12237-014-9928-9 |
MCCARTHY M J, CARINI S A, LIU Z F, et al, 2013. Oxygen consumption in the water column and sediments of the northern Gulf of Mexico hypoxic zone [J]. Estuarine, Coastal and Shelf Science, 123: 46-53.
|
MILLÁN-NÚÑEZ E, SIERACKI M E, MILLÁN-NÚÑEZ R, et al, 2004. Specific absorption coefficient and phytoplankton biomass in the southern region of the California Current[J]. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 51(6/7/8/9): 817-826. |
MOREL A, BRICAUD A, 1981. Theoretical results concerning light absorption in a discrete medium, and application to specific absorption of phytoplankton. Deep Sea Research Part A[J]. Oceanographic Research Papers, 28(11): 1375-1393. |
MORSE J W, ELDRIDGE P M, 2007. A non-steady state diagenetic model for changes in sediment biogeochemistry in response to seasonally hypoxic/anoxic conditions in the "dead zone" of the Louisiana shelf[J]. Marine Chemistry, 106(1/2): 239-255. |
OLSSON P, GRANÉLI E, CARLSSON P, et al, 1992. Structuring of a postspring phytoplankton community by manipulation of trophic interactions[J]. Journal of Experimental Marine Biology and Ecology, 158(2): 249-266. DOI:10.1016/0022-0981(92)90230-8 |
PARSONS T R, MAITA Y, LALLI C M, 1984. Fluorometric determination of chlorophylls [M]//PARSONS T R, MAITA Y, LALLI C M. A Manual of Chemical & Biological Methods for Seawater Analysis. New York: Pergamon: 107-109.
|
QIU Z F, SUN D Y, HU C M, et al, 2016. Variability of particle size distributions in the Bohai Sea and the Yellow Sea[J]. Remote Sensing, 8(11): 949. DOI:10.3390/rs8110949 |
RABALAIS N N, DÍAZ R J, LEVIN L A, et al, 2010. Dynamics and distribution of natural and human-caused hypoxia[J]. Biogeosciences, 7(2): 585-619. DOI:10.5194/bg-7-585-2010 |
RABALAIS N N, TURNER R E, GUPTA B K S, et al, 2007. Sediments tell the history of eutrophication and hypoxia in the northern gulf of Mexico[J]. Ecological Applications, 17(S5): S129-S143. |
REYNOLDS R A, STRAMSKI D, 2021. Variability in oceanic particle size distributions and estimation of size class contributions using a non-parametric approach[J]. Journal of Geophysical Research: Oceans, 126(12): e2021JC017946. DOI:10.1029/2021JC017946 |
RÖTTGERS R, GEHNKE S, 2012. Measurement of light absorption by aquatic particles: improvement of the quantitative filter technique by use of an integrating sphere approach[J]. Applied Optics, 51(9): 1336-1351. DOI:10.1364/AO.51.001336 |
SATHYENDRANATH S, COTA G, STUART V, et al, 2001. Remote sensing of phytoplankton pigments: a comparison of empirical and theoretical approaches[J]. International Journal of Remote Sensing, 22(2/3): 249-273. |
SATHYENDRANATH S, LAZZARA L, PRIEUR L, 1987. Variations in the spectral values of specific absorption of phytoplankton[J]. Limnology and Oceanography, 32(2): 403-415. DOI:10.4319/lo.1987.32.2.0403 |
SATHYENDRANATH S, PLATT T, 1988. The spectral irradiance field at the surface and in the interior of the ocean: a model for applications in oceanography and remote sensing[J]. Journal of Geophysical Research: Oceans, 93(C8): 9270-9280. DOI:10.1029/JC093iC08p09270 |
SONG G S, ZHAO L, CHAI F, et al, 2020. Summertime oxygen depletion and acidification in Bohai Sea, China[J]. Frontiers in Marine Science, 7: 252. DOI:10.3389/fmars.2020.00252 |
STRAMSKI D, SCIANDRA A, CLAUSTRE H, 2002. Effects of temperature, nitrogen, and light limitation on the optical properties of the marine diatom Thalassiosira pseudonana[J]. Limnology and Oceanography, 47(2): 392-403. DOI:10.4319/lo.2002.47.2.0392 |
STUART V, SATHYENDRANATH S, PLATT T, et al, 1998. Pigments and species composition of natural phytoplankton populations: effect on the absorption spectra[J]. Journal of Plankton Research, 20(2): 187-217. DOI:10.1093/plankt/20.2.187 |
SUN X R, SHEN F, BREWIN R J W, et al, 2022. Light absorption spectra of naturally mixed phytoplankton assemblages for retrieval of phytoplankton group composition in coastal oceans[J]. Limnology and Oceanography, 67(4): 946-961. DOI:10.1002/lno.12047 |
TASSAN S, FERRARI G M, 1995. An alternative approach to absorption measurements of aquatic particles retained on filters[J]. Limnology and Oceanography, 40(8): 1358-1368. DOI:10.4319/lo.1995.40.8.1358 |
TURNER A, MILLWARD G E, 2002. Suspended particles: their role in estuarine biogeochemical cycles[J]. Estuarine, Coastal and Shelf Science, 55(6): 857-883. DOI:10.1006/ecss.2002.1033 |
WEI H, ZHAO L, ZHANG H Y, et al, 2021. Summer hypoxia in Bohai Sea caused by changes in phytoplankton community[J]. Anthropocene Coasts, 4(1): 77-86. DOI:10.1139/anc-2020-0017 |
WOŹNIAK B, DERA J, KOBLENTZ-MISHKE O J, 1992. Modeling the relationship between primary production, optical properties, and nutrients in the sea [C]//Proceedings of SPIE 1750, Ocean Optics XI. San Diego: SPIE: 246-275.
|
ZHAI W D, ZHAO H D, ZHENG N, et al, 2012. Coastal acidification in summer bottom oxygen-depleted waters in northwestern-northern Bohai Sea from June to August in 2011[J]. Chinese Science Bulletin, 57(9): 1062-1068. DOI:10.1007/s11434-011-4949-2 |
ZHANG H Y, ZHAO L, SUN Y, et al, 2017. Contribution of sediment oxygen demand to hypoxia development off the Changjiang Estuary [J]. Estuarine, Coastal and Shelf Science, 192: 149-157.
|
ZHAO H D, KAO S J, ZHAI W D, et al, 2017. Effects of stratification, organic matter remineralization and bathymetry on summertime oxygen distribution in the Bohai Sea, China[J]. Continental Shelf Research, 134: 15-25. DOI:10.1016/j.csr.2016.12.004 |