 2024, Vol. 55
2024, Vol. 55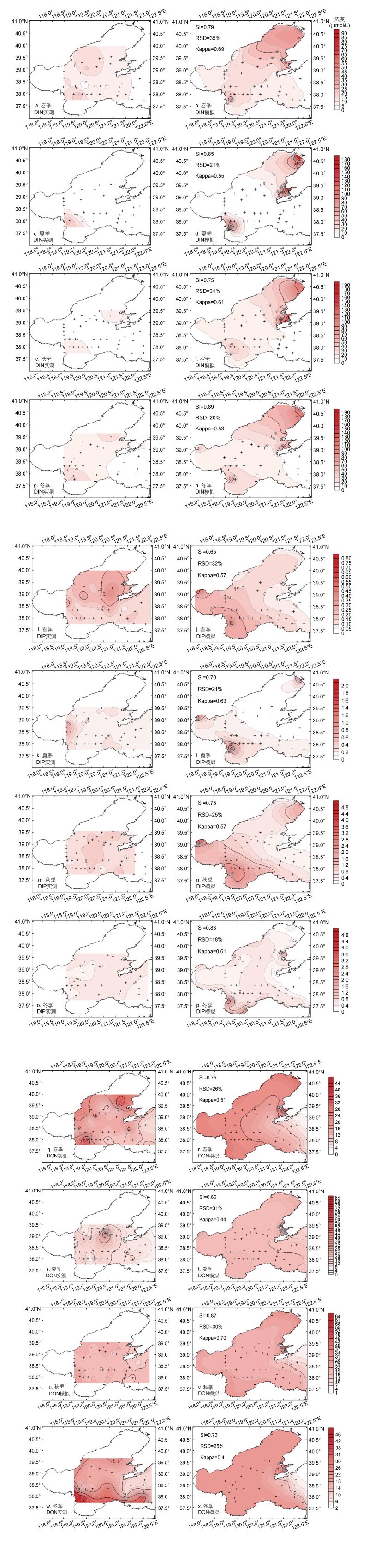


渤海氮磷营养盐和叶绿素浓度时空分布数值模拟与富营养化评估

http://dx.doi.org/10.11693/hyhz20230700141
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 赵宇, 李克强, 孙珊, 陈衎, 谭光深, 张娟, 王修林. 2024.
- ZHAO Yu, LI Ke-Qiang, SUN Shan, CHEN Kan, TAN Guang-Shen, ZHANG Juan, WANG Xiu-Lin. 2024.
- 渤海氮磷营养盐和叶绿素浓度时空分布数值模拟与富营养化评估
- NUMERICAL SIMULATION AND EUTROPHICATION ASSESSMENT OF SPATIOTEMPORAL DISTRIBUTION OF NITROGEN, PHOSPHORUS, AND CHLOROPHYLL CONCENTRATIONS IN THE BOHAI SEA
- 海洋与湖沼, 55(1): 118-134
- Oceanologia et Limnologia Sinica, 55(1): 118-134.
- http://dx.doi.org/10.11693/hyhz20230700141
文章历史
-
收稿日期:2023-07-08
收修改稿日期:2023-10-09
引用本文
赵宇, 李克强, 孙珊, 陈衎, 谭光深, 张娟, 王修林. 2024. 渤海氮磷营养盐和叶绿素浓度时空分布数值模拟与富营养化评估[J]. 海洋与湖沼, 55(1): 118-134

ZHAO Yu, LI Ke-Qiang, SUN Shan, CHEN Kan, TAN Guang-Shen, ZHANG Juan, WANG Xiu-Lin. 2024. NUMERICAL SIMULATION AND EUTROPHICATION ASSESSMENT OF SPATIOTEMPORAL DISTRIBUTION OF NITROGEN, PHOSPHORUS, AND CHLOROPHYLL CONCENTRATIONS IN THE BOHAI SEA[J]. Oceanologia et Limnologia Sinica, 55(1): 118-134.
渤海氮磷营养盐和叶绿素浓度时空分布数值模拟与富营养化评估
1. 中国海洋大学海洋化学理论与工程技术教育部重点实验室 中国海洋大学化学化工学院 山东青岛 266100;
2. 山东省海洋资源与环境研究院 山东烟台 264006
2. 山东省海洋资源与环境研究院 山东烟台 264006
摘要:当前渤海富营养化风险仍居高不下, 严重制约了环渤海社会经济可持续发展。开展近海富营养化评估与趋势分析是国家生态安全保障的需求, 其难点在于富营养化评价要素长期演变进程高质量数据的获取。基于HAMSOM海洋生态模型, 通过修正溶解有机氮(dissolved organic nitrogen, DON)的难/易降解组分降解动力学形式, 构建了适用于富营养化评估与趋势分析的渤海三维水动力/生物地球化学耦合模型。利用2019年渤海春、夏、秋、冬四个季节DON、溶解无机氮(dissolved inorganic nitrogen, DIN)、溶解无机磷(dissolved inorganicphosphorus, DIP)和叶绿素a (chlorophyll a, chl a)调查结果, 对模型进行了校正, 并利用1980年至2020年的长期调查结果进行了验证, 模拟结果与调查结果相比较, 在数值大小和变化趋势上均吻合较好, 相对标准偏差、相似性系数和Kappa系数分别为24%、0.77和0.60。利用模型模拟计算的DIN、DIP、DON和chl a长期演变进程数据, 计算了营养状态质量指数(nutritional quality index, NQI)。结果表明, 当前渤海富营养化状态整体上处于贫营养状态, 但在渤海湾、辽东湾和莱州湾湾底近岸海域处于富营养状态, 季节上5~10月份处于中等富营养化状态, 从长期变化趋势看, 渤海整体上富营养化状态趋于改善。与复合富营养化指数(compound eutrophication index, CEI)对比表明, 渤海富营养化评估按NQI计算结果与按CEI计算结果相当吻合, 相似性系数为0.83。文章建立的渤海富营养化评估方法具有较高可靠性, 可用于渤海富营养化评估。
关键词:渤海 生态模型 富营养化评估 数值模拟
NUMERICAL SIMULATION AND EUTROPHICATION ASSESSMENT OF SPATIOTEMPORAL DISTRIBUTION OF NITROGEN, PHOSPHORUS, AND CHLOROPHYLL CONCENTRATIONS IN THE BOHAI SEA
1. Key Laboratory of Marine Chemistry Theory and Engineering Technology, Ministry of Education, Ocean University of China, Qingdao 266100, China;
2. Shandong Institute of Marine Resources and Environment, Yantai 264006, China
2. Shandong Institute of Marine Resources and Environment, Yantai 264006, China
Abstract: At present, the risk of eutrophication in the Bohai Sea remains high, seriously constraining the sustainable development of the social and economic environment around the Bohai Sea. Conducting assessment and trend analysis of nearshore eutrophication is required for national ecological security. However, the difficulty lies in obtaining high-quality data on the long-term evolution process of eutrophication assessment elements. Based on the HAMSOM marine ecological model, we constructed a three-dimensional hydrodynamic/biogeochemical coupling model suitable for eutrophication assessment and conducted trend analysis for the Bohai Sea by modifying the degradation kinetics of dissolved organic nitrogen (DON) as a difficultly/easily degradable component. The model was calibrated using the survey results of DON, dissolved inorganic nitrogen (DIN), dissolved inorganic phosphorus (DIP), and chlorophyll a (chl a) in four seasons of 2019 in the Bohai Sea. The model was validated using long-term survey results from 1980 to 2020. Results show that the simulation results agree well with the survey results in terms of numerical magnitude and trend. The relative standard deviation, similarity coefficient, and Kappa coefficient are 24%, 0.77, and 0.60, respectively. The nutritional quality index (NQI) was calculated using long-term evolution process data of DIN, DIP, DON, and chl a simulated by the model. The results indicate that the current eutrophication status of the Bohai Sea is generally in oligotrophic state, while in eutrophic state in the coastal waters of the Bohai Bay, Liaodong Bay, and Laizhou Bay but in mesotrophic state from May to October. In long-term view, the overall eutrophication status of the Bohai Sea tended to improve. The compound eutrophication index (CEI) shows that the eutrophication assessment results calculated by CEI are quite consistent with those calculated by NQI, with a similarity coefficient of 0.83. The eutrophication assessment method established in this study has high reliability and can be used for eutrophication assessment in the Bohai Sea.
Key words:
the Bohai sea ecological model assessment of eutrophication numerical simulation
当前, 富营养化是海湾主要的生态环境问题。全球经济中心大多坐落在环湾地区, 随着陆源污染物入海排放等人类活动压力逐渐增大, 海湾富营养化评估和污染防治成为全球陆海环境综合治理的重点。自1960年代末特别是1972年联合国第一次人类环境会议以来, 美国、日本、欧盟诸国相继开始近海富营养化综合治理, 其中, 厘清水体富营养化状态与变化趋势是综合治理的关键, 获取营养盐和叶绿素等富营养化评估要素长期的演变进程高质量数据是前提和难点。在海洋调查监测数据部分年代缺失条件下, 数值模型模拟计算是重构营养盐和叶绿素等富营养化评估要素数据集的有效手段, 当前逐步发展建立了EFDC、WASP、FVCOM、ECOHAM、ERSEM、COHERENS等多种数值模型计算方法(任湘湘, 2011; 樊嘉蓉, 2022)。按照DPSIR理论等, 海湾营养盐、有机质、叶绿素等富营养化评估要素, 主要是环湾地区各种人类经济社会活动压力共同作用的结果。这不仅表现在陆源污染物入海排放等改变氮磷营养盐等化学变量的浓度、结构和组成, 也表现在围填海、海岸工程等通过改变流速、流向等水动力场物理变量, 间接影响营养盐、有机质、叶绿素等生地化变量的时空分布的过程。因此, 海湾富营养化评估要素的数值模拟需要物理、化学和生物多种变量的模拟计算, 其中生物生态过程是长时间的演变过程, 需要进行多变量长时序模拟计算。
海湾富营养化评估要素(营养盐、有机质、叶绿素等)的模拟计算, 相关研究始于20世纪40年代(房月英, 2008)。早期研究聚焦富营养盐、叶绿素等单变量季节或时空分布模拟, 至20世纪80年代(Skogen et al, 2000; Moll et al, 2003), 海洋三维水动力/生物地球化学模型模拟研究进一步完善和发展, 主要包括海湾富营养化要素控制过程及作用机制研究、模型逻辑架构构建及修正、参数率定、模拟计算等。海湾富营养化要素时空变化是水动力输运、生物地球化学迁移转化、浮游植物生长等过程综合作用的结果。对于海湾水动力输运过程研究, 通常采用现场观测结合数值模拟的方法研究流场中潮流和余流变化规律(李高阳, 2015)。对于营养盐生物地球化学迁移转化和浮游植物生长过程, 通常采用陆海同步调查、围隔生态系/培养瓶现场培养实验和数值模型模拟方法研究(Chen et al, 2022)。对于模型逻辑架构, 海湾水动力模型主要包括ROMS、FVCOM、POM、HAMSOM等, 目前广泛采用ROMS和FVCOM; 生物地球化学和生态动力学模型主要包括WASP、NPZD、ERSEM等(刘学海, 2009)。对于模型参数率定, 通常采用伴随矩阵法(吕咸青等, 2002)、模拟退火法(张艳杰等, 2006)、蒙特卡罗法(朱陆陆, 2014)等。对于模拟计算技术, 通常采用粒子追踪法(翁怡婵, 2009)、差分法(Kristek et al, 2010)和分步模拟计算方法(齐宁等, 2020)等。
最近, Gao等(2020)发展完善了海洋围隔生态系/实验瓶现场培养实验方法, 这不仅揭示了莱州湾浮游植物优势种与不同形态氮迁移转化过程之间的关系, 而且从动力学上量化了莱州湾几种典型浮游植物优势种演替的营养盐控制过程和作用机制。同时, Li等(2017)根据渤海三维水动力/生物地球化学耦合模型, 应用陆海水环境同步监测调查数据, 将渤海溶解无机氮(dissolved inorganic nitrogen, DIN)等浓度时空分布模拟计算结果的准确性平均提高约25%。这样, 围绕渤海富营养化评估要素的数值模拟计算, 针对渤海三维水动力/生物地球化学耦合模型在逻辑架构、过程耦合、参数地域化等存在的问题, 本文在改进渤海三维水动力/生物地球化学过程耦合模型架构基础上, 结合陆海同步调查监测数据校正、参数率定、多变量模拟计算等方法, 建立完善了渤海富营养化评估要素长期演变数据重构技术方法, 据此利用修订的营养状态质量指数(Nutritional Quality Index, NQI), 科学评估了渤海富营养化状态和变化趋势, 为进一步提高中国推行的渤海重点海域“深入打好污染防治攻坚战”行动计划的实施提供坚实的科技支撑。
1 研究方法海湾多变量模拟计算准确性不仅取决于模型的逻辑架构和监测数据的质量, 也取决于模型参数的灵敏度。这样, 海湾富营养化评估要素模拟计算主要包括三维水动力/生物地球化学耦合数值模型构建、参数率定、多变量模拟计算与准确性评价等步骤。
1.1 数值模型构建王强(2004)和Dai等(2015)在Hamburg Shelf Ocean Model (HAMSOM)(Backhaus et al, 1983)基础上建立了渤海三维水动力/生物地球化学耦合模型, 本文在此基础上, 对相关生物地球化学模块进行修正。模型中入海口相关数据参见戴爱泉(2015)研究结果。
根据溶解有机氮(dissolved organic nitrogen, DON)的现场培养实验结果(戴爱泉, 2015; Li et al, 2017), 不同来源的DON结构组成和降解速率存在显著差异。本文按照污水处理厂和河流不同来源DON的降解速率差异性, 将生物地球化学过程模型中DON状态变量分为污水处理厂输入生物易降解DON(BDON1)、河流输入生物易降解DON(BDON2)和生物难降解DON(RDON), 方程修正如下:
 (1)
(1) (2)
(2)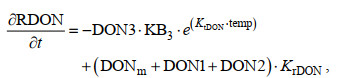 (3)
(3)式中, DON1为陆源污水处理厂输入的生物易降解DON浓度; DON2为河流输入的生物易降解DON浓度; DON3为各排污口排入的难降解DON浓度; DONm为海源DON浓度; KrDON为难降解DON组分占比; KB1、KB2、KB3分别为DON1、DON2、DON3的降解速率常数; KtDON为溶解有机氮微生物降解温度效应系数; temp为环境温度。
结合生物地球化学模块其他状态变量迁移转化过程, 建立的模型主要框架及过程如图 1所示。模型主要包括浮游植物、浮游动物、溶解无机态营养盐、溶解有机态营养盐、生物碎屑5个子模块。共包含DIN、陆源生物易降解DON (BDON1, BDON2)、海源DON (BDONm)、生物难降解DON(RDON)、无机磷(DIP)、有机磷(DOP)、浮游植物(PHYTO)、浮游动物(ZOO)、碎屑(DPT)10个状态变量。物质流统一用氮元素(单位: mmol/m3)表示, 浮游植物生物量可表示为活性颗粒态氮(PPON), chl a浓度通过浮游植物叶绿素氮比值关系进行换算(表 1)。主要的生物地球化学过程有: 浮游植物生长、死亡和营养盐吸收分泌等过程; 浮游动物生长、死亡和营养盐释放等过程; 无机态和有机态营养盐吸收和转化过程; 碎屑矿化和沉降过程等。相关生物地球化学过程动力学方程详见附表Ⅰ。
 |
| 图 1 渤海三维水动力/生物地球化学耦合模型架构示意图 Fig. 1 Schematic architecture of three-dimensional hydrodynamic/biogeochemical multi-process coupled model for the Bohai Sea 注: (1)浮游植物吸收过程; (2)浮游植物呼吸过程; (3)有机态营养盐经微生物降解为无机态营养盐过程; (4)浮游植物分泌过程; (5)浮游动物分泌过程; (6)浮游动物摄食过程; (7)浮游生物死亡过程; (8)碎屑经细菌分解为有机态营养盐过程; (9)沉降过程; (10)再矿化过程。DIN: 溶解无机氮; DIP: 溶解无机磷; BDON1、BDON2: 陆源污水处理厂和河流输入生物易降解溶解有机氮: BDONm: 海源溶解有机氮; RDON: 生物难降解溶解有机氮; PHYTO: 浮游植物; ZOO: 浮游动物; DPT: 生物碎屑 |
表 1 模型动力学过程所用参数意义及取值 Tab. 1 Significance and value of parameters used in model dynamics process
| 符号 | 意义 | 值 | 单位 | 来源 |
| parf | 光衰减系数 | 0.6 | 1/m | Fasham et al, 1990 |
| par0 | 海水自身消光系数 | 0.8 | 1/m | Fasham et al, 1990 |
| par1 | 浮游植物自遮光系数 | 0.0088 | 1/(m·mg chla) | Riley, 1956 |
| par2 | 浮游植物自遮光系数 | 0.054 | 1/(m·mg chla2/3) | Riley, 1956 |
| parm | 浮游植物生长最佳光合有效辐射 | 90 | W/m2 | 俞光耀等, 1999 |
| KPPTG | 浮游植物生长速率常数 | 2.8 | 1/d | Eppley, 1972 |
| KPPTD | 浮游植物死亡速率常数 | 0.05 | 1/d | Eppley, 1972 |
| KZPTG | 浮游动物生长速率常数 | 0.4 | 1/d | Jørgensen et al, 1991 |
| KZPTD | 浮游动物死亡速率常数 | 0.05 | 1/d | Jørgensen et al, 1991 |
| KZPTN | 浮游动物代谢速率常数 | 0.2 | 1/d | Wen et al, 1994 |
| KZPTF | 浮游动物被捕食速率常数 | 0.1 | 1/d | Li et al, 2017 |
| KDPTZ | 浮游动物摄食生物碎屑速率常数 | 0.6 | 1/d | Fasham et al, 1990 |
| KrDON | 难降解DON组分占比 | 0.0002 | — | — |
| KB1 | 污水处理厂输入半易降解DON的降解速率常数 | 0.07 | 1/d | Li et al, 2017 * |
| KB2 | 河流输入半易降解DON的降解速率常数 | 0.1 | 1/d | Li et al, 2017 * |
| KB3 | 各排污口输入难降解DON的降解速率常数 | 0.001 | 1/d | Li et al, 2017 * |
| rchlpn | 浮游植物叶绿素氮比值 | 1.2 | mg chla/mmolN | 俞光耀等, 1999 |
| rpptz | 浮游动物捕食浮游植物同化系数 | 0.8 | — | Chapelle et al, 1994 |
| rdptz | 浮游动物捕食生物碎屑同化系数 | 0.7 | — | Savchuk et al, 2002 |
| rpnp | 氮磷比 | 16 | — | Redfield et al, 1963 |
| KsDIN | 浮游植物吸收DIN的半饱和常数 | 8.5 | mmol N/m3 | 张蕾等, 2002 |
| KsDIP | 浮游植物吸收PO4-P的半饱和常数 | 0.6 | mmol P/m3 | 张蕾等, 2002 |
| KsDPT | 生物碎屑被摄食的半饱和常数 | 0.7 | mmol N/m3 | Fasham et al, 1990 |
| KsPPT | 浮游植物被摄食的半饱和常数 | 0.6 | mmol N/m3 | Radach et al, 1993 |
| KcPPT | 浮游植物被摄食的最低阈值浓度 | 0.12 | mmol N/m3 | Radach et al, 1993 |
| KtDON | DON降解温度效应系数 | 0.05 | 1/℃ | Chapelle et al, 1994 * |
| KtPPTG | 浮游植物生长温度效应系数 | 0.065 | 1/℃ | Eppley, 1972 |
| KtPPTD | 浮游植物死亡温度效应系数 | 0.05 | 1/℃ | Eppley, 1972 * |
| KtZPTD | 浮游动物死亡温度效应系数 | 0.05 | 1/℃ | Jørgensen et al, 1991 |
| KtZPTN | 浮游动物代谢温度效应系数 | 0.027 | 1/℃ | Jørgensen et al, 1991 |
| vDPT | 碎屑沉降速率 | 0.025 | m/d | Fasham et al, 1990 |
| 注: 上标*表示在初始值基础上进行了校正 | ||||
参数设置在主要参考文献值基础上, 采用渤海莱州湾海域现场培养实验结果(李克强等, 2007; Li et al, 2009, 2017), 利用DIN、DIP、DON、chl a浓度季节变化平面分布模拟结果与实测结果对比进行校正, 利用年际变化模拟结果与实测结果对比进行验证, 模型参数结果如表 1所示。
1.2.2 数据来源模型校验数据主要取自中国近岸海域环境质量公报(报告书)、中国海洋环境质量(状况)公报、北海区海洋环境公报、中国海洋生态环境状况公报和文献结果, 详见表 2。其中, 2019年氮磷营养盐和chl a浓度来源于国家自然科学基金委渤黄海共享航次(航次编号: NORC2018-01; NORC2019-01)调查结果。
表 2 模型模拟计算结果验证数据来源 Tab. 2 Data source for validation on the modelling results
| 集群 | 要素: 单位 | 数据来源 |
| 生物地球化学要素 | 溶解无机氮浓度(CDIN): μmol/L 溶解无机磷浓度(CDIP): μmol/L |
王修林等, 2006 崔毅等, 1996 王燕等, 2021 Li et al, 2023 |
| 溶解有机氮浓度(CDON): μmol/L | 王婷, 2009 李志林等, 2015 唐永等, 2017 |
|
| 叶绿素浓度(chl a): μg/L | 朱明远等, 1982 吕培顶等, 1984 吕瑞华等, 1999 赵骞等, 2004 田洪阵等, 2019 孟庆辉等, 2022 |
|
| 污染物输入通量 | 溶解无机氮入海通量(FDIN): t 溶解无机磷入海通量(FDIP): t 溶解有机氮入海通量(FDON): t |
王修林等, 2006 2009年渤海海洋环境公报(国家海洋局北海分局, 2010) 北海区海洋环境公报, 2010—2017(国家海洋局北海分局, 2011—2018) 中国近岸海域环境质量公报(报告书), 2001—2016(中国环境监测总站等, 2002—2017) 中国海洋环境质量(状况)公报, 2000-2017(国家海洋局, 2001—2018) 中国海洋生态环境状况公报, 2018—2020(中华人民共和国生态环境部, 2019—2021) |
本文选用余弦相似性系数(similarity index, SI; 方程4)和相对标准偏差(relative standard deviation, RSD; 方程5)分别评价模拟值和实测值平面分布趋势相似性以及模拟值偏离实测值的程度(戴爱泉, 2015), 从而综合定量评价模型模拟结果的准确性:
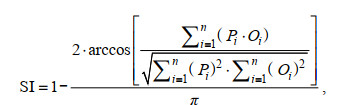 (4)
(4) (5)
(5)其中, Oi和Pi分别表示第i个水质点上的实测值和模拟值; n表示监测站位数量。
针对渤海DIN、DIP、DON与chl a浓度分布场, 同时使用Kappa系数评价模型模拟计算准确性。采用Cohen提出的Kappa系数评价分类标准(Cicchetti et al, 1990; Feinstein et al, 1990)(表 3)。
表 3 Kappa系数评价分类标准 Tab. 3 Classification criteria for Kappa coefficient evaluation
| Kappa系数 | 0.00~0.20 | 0.21~0.40 | 0.41~0.60 | 0.61~0.80 | 0.81~1.00 |
| 一致性程度 | 微弱(slight) | 一般(fair) | 中等(moderate) | 显著(substantial) | 极佳(almost perfect) |
渤海海水中溶解有机氮(DON)的浓度近年来持续升高, 某些海域已成为主要的氮形态(李志林等, 2015)。在富营养化评价方法中, 用DON代替化学需氧量(COD)能更好地评估富营养化状态。本文应用营养状态质量指数(NQI)法(陈于望, 1987), 采用DON代替COD予以修订, 计算公式如下:
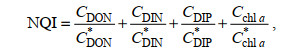 (6)
(6)式中, C为下标各要素DON、DIN、DIP与chl a的浓度值, 上标*表示各要素DON、DIN、DIP与chl a的营养状态评价标准。其中, chl a的营养状态评价标准采用赤潮发生叶绿素浓度阈值, 其他评价要素参照海水水质标准(GB 3097-1997), DON采用DIN标准值, 各要素营养状态评价标准分别取值为CDON*= 35.71μmol/L, CDIN*=35.71μmol/L, CDIP*=1.45μmol/L, Cchl a*=5μg/L。这样, 通过各要素浓度与标准的比值可消掉量纲。NQI>3为富营养状态, 3≥NQI≥2为中等营养状态, NQI<2为贫营养状态, NQI指数越高富营养化程度则越高。
2 结果与讨论 2.1 渤海氮、磷和chl a浓度时空变化模拟计算结果 2.1.1 DIN、DIP和DON平面分布应用构建完善的三维水动力/生物地球化学耦合数值模型, 模拟计算了2019年DIN、DIP、DON春夏秋冬四个季节的浓度平面分布(图 2)。经模型参数校正, 再与调查站位上的实测结果对比, DIN、DIP、DON浓度模拟结果在数值大小及变化趋势上均具有较高的一致性, 三者SI平均值为0.77±0.08, RSD平均值为26%±5%, Kappa系数平均值为0.57±0.08。
结果表明, 模拟计算值再现了渤海的DIN、DIP与DON浓度平面分布春夏秋冬的季节变化特征。在春、夏两季, DIN浓度因浮游植物生长被消耗利用, 渤海湾与辽东湾南部浓度都比较低, 莱州湾西部受黄河径流输入的影响, 成为夏季渤海DIN浓度最高的海区(图 2b, 2d); 秋季渤海DIN浓度高值区不断扩大, 渤海中部浓度相对较低, 但也高于夏季水平; 冬季渤海DIN浓度达到最高峰, 浓度高值区较冬季继续扩大, 渤海中部平均浓度达到秋季的两倍水平, 仅有渤海海峡东部浓度较低(图 2h)。由于浮游植物在春、夏季生长的消耗, 海水中的DIP浓度较低(图 2j, 2l); 秋季渤海湾南岸、莱州湾以及辽东湾北岸DIP浓度较高, 渤海中部浓度较低, 夏秋季是我国北方降雨期, 三大湾沿岸DIP浓度上升与陆地径流输入有关, 从而形成了由沿岸高值区向渤海中部逐渐递减的趋势(图 2l, 2n)(王燕等, 2021); 冬季渤海中DIP含量上升, 主要因为冬季时浅海风力增大, 导致跃层失去了稳定性, 从而促使海水底层颗粒的有机磷分解所产生的DIP随着垂直混合被带到了表层(图 2p)(黄爽, 2012)。在春、秋和冬季, DON高值区主要集中于莱州湾西部、渤海湾、秦皇岛近岸、辽东湾北部及金州湾沿岸地区(图 2r, 2v, 2x); 而夏季DON受高值区出现在渤海中部(图 2t), 模型没有很好的模拟再现这一特征, 这与DON等有机质受浮游植物和微生物调控有关, 由于浮游植物夏季较高的生物量, DON的源汇过程较为复杂, 模型模拟很难实现精准刻画, 但在调查站位上与调查监测结果比较, 31%的RSD值还是符合海上调查监测平均误差范围(约30%)的。
在季节变化上, 2019年渤海夏季DIN、DIP浓度显著低于其他季节, 而秋、冬季DON浓度高于其他季节(图 2), 这与王婷(2009)在2002~2004年的研究结果在变化规律上基本一致, 构建的模型很好的体现了黄河调水调沙引起的营养盐输入通量变化对DIN和DIP浓度平面分布产生的影响(王婷, 2009; Ding et al, 2020)。
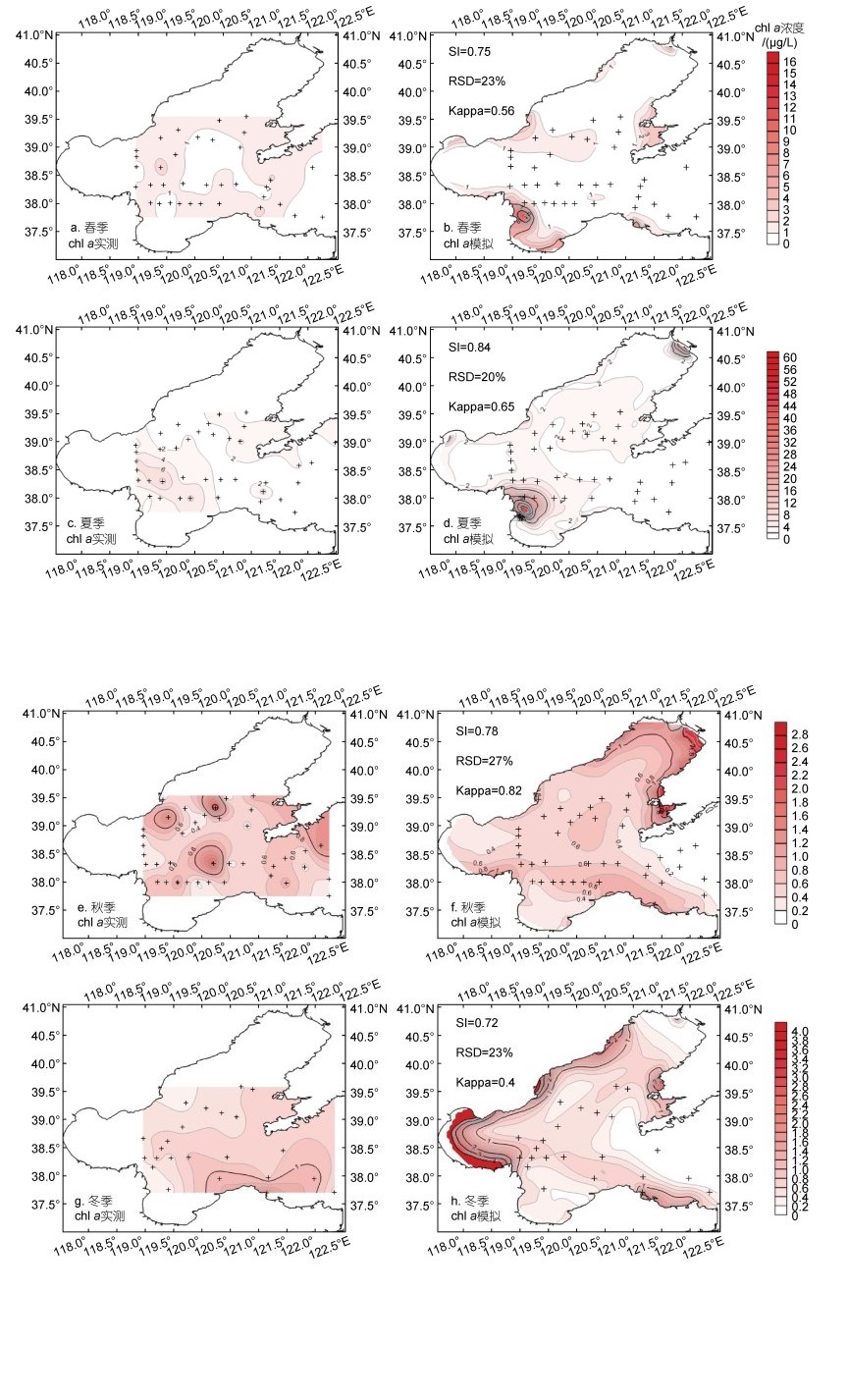
2.1.2 chl a浓度平面分布应用模型模拟计算了2019年chl a春夏秋冬四个季节的浓度平面分布(图 3)。在调查站位上与调查监测结果对比, chl a浓度模拟结果在数值大小及变化趋势上均具有较高的一致性, SI平均值为0.77±0.04, RSD平均值为23%±2%, Kappa系数平均值为0.63±0.13。
渤海的chl a浓度平面分布模拟计算结果体现了浮游植物春夏秋冬的季节变化特征。春季chl a浓度的高值区出现在黄河口附近海域、莱州湾南部、渤海湾与辽东湾北部、秦皇岛外海与金州湾, 渤海中央以及辽东湾南部海区为低值区(图 3b); 夏季chl a浓度高值区主要集中于黄河与辽河入海口附近海域, 中央海区浓度较春季增加一倍(图 3d); 秋季由于光照、温度等因素影响, chl a浓度急剧下降, 仅在辽东湾沿岸出现高值区(图 3f); 冬季整个渤海的表层chl a浓度继续下降, 只在渤海湾沿岸与秦皇岛沿岸地区浓度较高(图 3h)。
在季节变化上, 2019年渤海夏季chl a浓度显著高于其他季节, chl a与DIN和DIP浓度的季节性分布趋势呈负相关, 与DON浓度季节性分布趋势呈正相关(图 3)。主要的浓度高值区分布在黄河口临近海域, 构建的模型很好的体现了黄河调水调沙引起的营养盐输入通量变化对浮游植物生物量平面分布产生的影响(Ding et al, 2020)。
综上, DIN、DIP、DON与chl a浓度模拟值与实测值的相对标准偏差、相似性系数和Kappa系数平均值分别为24%、0.77和0.60, 说明通过该模型能够较真实的反映渤海海域DIN、DIP、DON与chl a浓度分布情况, 可以用于渤海DIN、DIP、DON与chl a浓度长期变化模拟计算和数据重构。
2.1.3 年际变化以1980年至2020年部分年代调查监测的DIN、DIP、DON和chl a浓度年际变化对模型进行验证。图 4中模拟值为模型对全渤海逐月模拟结果的平均值, 监测值为对应某一月份的所有监测值的平均值。模拟结果与调查监测结果比较表明, 模拟结果在数值大小与变化趋势上均能够很好的再现渤海近40年营养盐和chl a的年际变化, DIN、DIP、DON和chl a年际变化模拟结果与调查结果的RSD平均值小于20%, SI平均值高于0.80(图 4)。
 |
| 图 4 1980~2020年逐月DIN、DIP、DON、chl a浓度的模拟结果与观测资料的对比 Fig. 4 Comparison of simulated and observed data of DIN, DIP, DON, and chl a concentrations month by month from 1980 to 2020 注: a: DIN; b: DIP; c: DON; d: chl a |
模拟结果表明, 20世纪80年代至90年代中期DIN浓度显著增加, 1995年冬季DIN浓度达到最大值(24.4 μmol/L), 在1996年至2004年间, DIN浓度逐渐下降, 在2005年至2013年期间, DIN浓度再次呈现上升趋势, 近几年DIN浓度又呈下降趋势(图 4a), 构建的模型能很好地再现这两次DIN浓度峰值变化(王修林等, 2006; Wang et al, 2019)。分析表明, 农业化肥使用、污水排放等人类活动导致总氮河流输入和大气沉降通量增加, 是造成渤海DIN浓度升高的主要原因, 自2002年起水利部黄河委员会于夏季实施的调水调沙工程, 是导致渤海夏秋季营养盐输入和浓度增加的原因(Ding et al, 2020)。DIP浓度在20世纪80年代至90年代初逐渐下降, 在1992年夏季达到最小值(0.1 μmol/L), 然后增加至90年代后期, 在1998年冬季DIP浓度达到最大值(1.1 μmol/L), 之后呈下降趋势(图 4b), 模拟结果在年际变化和季节变化上均与文献结果基本一致(Ning et al, 2010; Xin et al, 2019)。20世纪90年代后期DIP浓度的下降, 与含磷洗涤剂的禁用有关。在20世纪80年代到90年代初DON浓度总体处于缓慢上升趋势, 直至1994年浓度有所下降; 21世纪10年代末有所上升, 在2014年又降低, 之后呈上升趋势(图 4c), 这与陆源长期输入变化与海源浮游植物、微生物长期演变过程有关(唐永等, 2017)。
Chl a浓度自20世纪80年代至21世纪初呈波动缓慢升高趋势, 在年际变化上与营养盐浓度, 特别是与DIN浓度变化呈现很好的相关性, 这与孟庆辉等(2022)的研究结果一致。Chl a浓度季节变化主要是夏季峰(图 4d), 与DIN浓度呈反向变化关系, 这与夏季藻华对营养物质的消耗和冬季浮游植物死亡营养盐恢复的生态学规律一致; 与DON浓度呈复杂源汇变化关系, 说明DON浓度主要受陆源和海源的影响, 也是支持浮游植物生长的重要物质(唐永等, 2017)。
2.2 模拟结果准确性评价为验证模型改进后对模拟计算结果准确性的提高程度, 将改进前后的模拟计算量化验证结果进行比较, 结果如图 5所示。氮、磷营养盐与chl a浓度的相似性系数平均值分别提高了0.07和0.13, Kappa系数分别提高了0.09和0.10, 说明改进后的模型对氮、磷营养盐与chl a浓度的变化趋势的模拟要显著比模型改进前与实测结果更加吻合。二者的相对标准偏差也均有明显降低, 氮、磷营养盐与chl a浓度分别降低了约11%和6%, 说明在数值大小上, 模型改进后的结果与实测结果也更为接近。综上所述, 通过参数优化和优质数据选取对模型进行改进, 提高了模型模拟计算的准确性。
 |
图 5 模型改进前后渤海氮、磷营养盐与chl a浓度模拟计算评价结果的Taylor图 Fig. 5 The Taylor diagram of the simulation results of N-P nutrients and chl a concentrations in Bohai Sea before and after model improvement 注:  分别为氮、磷营养盐改进前后的结果; 分别为氮、磷营养盐改进前后的结果;  为chl a浓度改进前后的结果 为chl a浓度改进前后的结果 |
渤海富营养化状态指数(NQI)年际变化计算结果表明, 自1980年至2020年, NQI整体呈先上升后下降的倒“U”型变化特征(图 6)。其中, 1980年至1995年, NQI上升达到极大值2.27, 1995年至2013年NQI缓慢下降, 2013年至2020年迅速下降; 从数值上看, 1993年至2017年, NQI基本都大于2, 富营养状态处于中等水平, 当前处于贫富营养状态。利用渤海复合富营养化指数(CEI)(Lin et al, 2020)年际变化计算结果进行后验, 结果表明, NQI与CEI在变化趋势上相当一致(SI=0.92) (图 6)。
 |
| 图 6 1980至2020年渤海复合富营养化指数(CEI)与富营养化状态(NQI)年际变化图 Fig. 6 Interannual changes of Bohai composite eutrophication index (CEI) and eutrophication status (NQI) from 1980 to 2020 注: 复合富营养化指数(CEI)结果引自(Lin et al, 2020) |
在渤海全海域富营养化状态季节变化规律上, NQI呈倒“U”型变化特征, 夏季富营养化程度最高; NQI在5~10月份超过2.0, 处于中等富营养状态。利用渤海复合富营养化指数(CEI)(Lin et al, 2020)季节变化结果对比后验, NQI与CEI在变化规律上也较为一致(SI=0.74)(图 7)。
 |
| 图 7 渤海复合富营养化指数(CEI)与富营养化状态(NQI)季节变化图 Fig. 7 Seasonal change in composite eutrophication index (CEI) and eutrophication status (NQI) in Bohai Sea 注: 复合富营养化指数(CEI)结果引自Lin等(2020) |
渤海富营养化状态指数(NQI)平面分布模拟计算结果如图 8所示。渤海富营养化状态平面分布总体呈现近岸较高、中部较低的趋势, 这与Lin等(2020)的研究结果一致。近岸较高的富营养化状态主要与人类活动有关, 且海湾近岸地区水体交换能力差, 二者综合导致近岸海域富营养化程度显著高于中部海域。在季节变化上, 富营养化状态与chl a浓度模拟结果(图 3)变化规律基本一致, 即随chl a浓度春夏季升高, 富营养化水平升高, 随chl a浓度秋冬季下降, 富营养化水平下降, 同时还受海洋中的环境因子(光照、温度、营养盐、混合层深度等)季节变化调控。
 |
| 图 8 2019年渤海富营养化状态(NQI)平面分布图 Fig. 8 Horizontal distribution of eutrophication status (NQI) in 2019 in the Bohai Sea 注: 黄、绿和蓝色分别表示富、中等和贫营养状态 |
本文通过改进生物地球化学模型架构, 建立完善了用于重构富营养化评价要素长期演变数据的渤海三维水动力/生物地球化学耦合数值模型。利用构建的模型成功模拟了渤海近四十年来DIN、DIP、DON与chl a浓度时空分布[RSD(相对标准偏差平均值)为24%, SI(余弦相似性系数平均值)为0.77, Kappa(Kappa系数平均值)为0.60], 说明模型相关动力学方程与参数能够反映渤海海域的地域化特征。应用模型模拟结果计算了营养状态质量指数(NQI), 与复合富营养化指数(CEI)具有较好的一致性(SI=0.83), 说明该模型能够反映渤海富营养化状况。
本文构建的三维水动力/生物地球化学耦合模型, 对富营养化状态的模拟计算结果能够较真实的反映渤海海域富营养化状态的时空变化情况, 建立的富营养化评估方法具有较高可靠性, 可以用于渤海富营养化评估, 并支撑渤海富营养化综合治理。
附表Ⅰ 生物地球化学过程生态动力学方程 AppendixⅠ Ecological dynamic equations Ⅰ discribing biogeochemistry process
| 方程 | 释义 |
 |
浮游植物的时间变化速率 |
| 光合作用下浮游植物渗出物的比例 | |
| 浮游植物光合作用速率 | |
| 浮游植物死亡速率 | |
| 浮游动物对浮游植物的捕食速率 | |
| 光对浮游植物的影响 | |
| 浮游动物的时间变化速率 | |
| 浮游动物对碎屑的捕食速率 | |
| 浮游动物死亡速率 | |
| 浮游动物排泄速率 | |
| 浮游动物被捕食速率 | |
| DIN的时间变化速率 | |
| DONm的降解速率 | |
| DIP的时间变化速率 | |
| DOPm的降解速率 | |
| DOP的时间变化速率 | |
| DPTm的降解速率 | |
| DPT的时间变化速率 | |
| 碎屑沉降速率 | |
| 注: par、temp为光照和温度, t、Hs为时间和水深, P、Z、D、DIN、DIP、DOPm分别为浮游植物、浮游动物、碎屑、溶解无机氮、溶解无机磷、海源溶解有机磷的状态变量浓度, Frpptn为光合作用下浮游植物渗出物的比例, FPPTG为浮游植物光合作用速率, FPPTD为浮游植物死亡速率, FZPTG为浮游动物对浮游植物的捕食速率, parm为浮游植物生长最佳光合有效辐射, KPPTG为浮游植物生长速率常数, KtPPTG为浮游植物生长温度效应系数, KsDIN为浮游植物吸收DIN的半饱和常数, KsDIP为浮游植物吸收PO4-P的半饱和常数, KPPTD为浮游植物死亡速率常数, KtPPTD为浮游植物死亡温度效应系数, KZPTG为浮游动物生长速率常数, KcPPT为浮游植物被摄食的最低阈值浓度, KsPPT为浮游植物被摄食的半饱和常数, parf为光衰减系数, par0为海水自身消光系数, par1、par2为浮游植物自遮光系数, rchlpn为浮游植物叶绿素氮比值, rpptz为浮游动物捕食浮游植物同化系数, rdptz为浮游动物捕食生物碎屑同化系数, FDPTZ为浮游动物对碎屑的捕食速率, FZPTD为浮游动物死亡速率, FZPTN为浮游动物排泄速率, FZPTF为浮游动物被捕食速率, rzptn浮游动物代谢同化系数, FDONm为DONm的降解速率, KRDON为难降解DON组分占比, KDONm为DONm的降解速率常数, KtDONm为DONm降解温度效应系数, FDOPm为DOPm的降解速率, rpnp为氮磷比, KDOPm为DOPm的降解速率常数, KtDOPm为DOPm降解温度效应系数, FDPTm为DPTm的降解速率, KDPTm为DPTm的降解速率常数, KtDPTm为DPTm降解温度效应系数, rzptf为浮游动物被捕食同化系数, FDPTB为碎屑沉降速率, vDPT为碎屑沉降速率 | |
参考文献
王修林, 李克强 , 2006. 渤海主要化学污染物海洋环境容量[M]. 北京: 科学出版社.
|
王强, 2004. 渤海环流的季节变化及浮游生态动力学模拟[D]. 青岛: 中国海洋大学.
|
王婷, 2009. 2002-2004年及调水调沙期间黄河下游营养盐的变化特征[D]. 青岛: 中国海洋大学.
|
王燕, 姚振童, 祝艳君, 等, 2021. 2019年渤海氮磷营养盐季节变化及富营养化状况[J]. 海洋环境科学, 40(6): 915-921. |
中华人民共和国生态环境部, 2019-2021. 2018-2020年中国海洋生态环境状况公报[R]. 北京: 中华人民共和国生态环境部.
|
中国环境监测总站和近岸海域环境监测中心站, 2002-2017. 中国近岸海域环境质量公报(报告书)(2001-2016年)[R]. 北京: 国家环境保护总局.
|
田洪阵, 刘沁萍, GOES J I, 等, 2019. 近20年渤海叶绿素a浓度时空变化[J]. 海洋学报, 41(8): 131-140. DOI:10.3969/j.issn.0253-4193.2019.08.015 |
吕咸青, 方国洪, 2002. 渤海开边界潮汐的伴随法反演[J]. 海洋与湖沼, 33(2): 113-120. DOI:10.3321/j.issn:0029-814X.2002.02.001 |
吕培顶, 费尊乐, 毛兴华, 等, 1984. 渤海水域叶绿素a的分布及初级生产力的估算[J]. 海洋学报, 6(1): 90-98. |
吕瑞华, 夏滨, 李宝华, 等, 1999. 渤海水域初级生产力10年间的变化[J]. 黄渤海海洋, 17(3): 80-86. |
朱陆陆, 2014. 蒙特卡洛方法及应用[D]. 武汉: 华中师范大学.
|
朱明远, 王艳香, 李宝华, 1982. 秋季渤海水域中叶绿素含量的测定及初级生产力的估算[J]. 海洋湖沼通报, (3): 42-46. |
任湘湘, 2011. COHERENS模式在渤海海洋环境模拟中的初步应用[D]. 北京: 国家海洋环境预报研究中心.
|
刘学海, 2009. 南黄海及养殖功能海域生态动力学模型研究[D]. 青岛: 中国海洋大学.
|
齐宁, 陈国彬, 李振亮, 等, 2020. 基于分步算法的裂缝性碳酸盐岩油藏大尺度酸化数值模拟[J]. 石油学报, 41(3): 348-362, 371. |
李志林, 石晓勇, 张传松, 2015. 春季黄渤海海水中尿素分布特征及溶解态氮的组成[J]. 环境科学, 36(11): 3999-4004. |
李克强, 王修林, 韩秀荣, 等, 2007. 莱州湾围隔浮游生态系统氮、磷营养盐迁移-转化模型研究[J]. 中国海洋大学学报, 37(6): 987-994. |
李高阳, 2015. 基于MIKE的渤海水动力数值模拟[J]. 中国水运, 15(6): 60-61. |
张艳杰, 郭建青, 王洪胜, 2006. 模拟退火算法在确定河流水质参数中的应用[J]. 中国农村水利水电, (1): 38-41. DOI:10.3969/j.issn.1007-2284.2006.01.015 |
张蕾, 王修林, 韩秀荣, 等, 2002. 石油烃污染物对海洋浮游植物生长的影响—实验与模型[J]. 中国海洋大学学报, 32(5): 804-810. |
陈于望, 1987. 厦门港海域营养状况的分析[J]. 海洋环境科学, 6(3): 14-19. |
国家海洋局, 2001-2018. 2000-2017年中国海洋环境质量(状况)公报[R]. 北京: 国家海洋局.
|
国家海洋局北海分局, 2010. 2009年渤海海洋环境公报[R]. 青岛: 国家海洋局北海分局.
|
国家海洋局北海分局, 2011-2018. 2010-2017年北海区海洋环境公报[R]. 青岛: 国家海洋局北海分局.
|
房月英, 2008. 三都湾赤潮监控区海水富营养化与赤潮发生的关系研究[D]. 福州: 福建师范大学.
|
孟庆辉, 王林, 陈艳拢, 等, 2022. 2002-2021年渤海叶绿素a浓度变化及环境响应分析[J]. 中国环境监测, 38(6): 228-236. |
赵骞, 田纪伟, 赵仕兰, 等, 2004. 渤海冬夏季营养盐和叶绿素a的分布特征[J]. 海洋科学, 28(4): 34-39. DOI:10.3969/j.issn.1000-3096.2004.04.008 |
俞光耀, 吴增茂, 张志南, 等, 1999. 胶州湾北部水层生态动力学模型与模拟Ⅰ. 胶州湾北部水层生态动力学模型[J]. 青岛海洋大学学报, 29(3): 421-428. |
翁怡婵, 杨金湘, 江毓武, 2009. 台湾海峡漂移物运动轨迹的数值模拟[J]. 厦门大学学报(自然科学版), 48(3): 446-449. DOI:10.3321/j.issn:0438-0479.2009.03.030 |
唐永, 孙语嫣, 石晓勇, 等, 2017. 黄渤海海域秋季营养盐及有色溶解有机物分布特征[J]. 环境科学, 38(11): 4501-4512. |
黄爽, 2012. 东海赤潮高发区颗粒有机物的来源、分布、分解及其环境效应[D]. 青岛: 中国海洋大学.
|
崔毅, 陈碧鹃, 任胜民, 等, 1996. 渤海水域生物理化环境现状研究[J]. 中国水产科学, 3(2): 1-12. |
樊嘉蓉, 2022. 基于EFDC和WASP耦合模型的水库水质数值模拟研究[J]. 湖南水利水电, (3): 78-80. |
戴爱泉, 2015. 渤海陆源TN和COD污染物分配容量指标体系计算方法及应用——环渤海经济圈356个县级行政区分配容量计算及35个地级行政区差别化减排措施构建[D]. 青岛: 中国海洋大学.
|
BACKHAUS J O, 1983. A semi-implicit scheme for the shallow water equations for application to shelf sea modelling[J]. Continental Shelf Research, 2(4): 243-254. DOI:10.1016/0278-4343(82)90020-6 |
CHAPELLE A, LAZURE P, MÉNESGUEN A, 1994. Modelling eutrophication events in a coastal ecosystem. Sensitivity analysis[J]. Estuarine, Coastal and Shelf Science, 39(6): 529-548. DOI:10.1016/S0272-7714(06)80008-9 |
CHEN K, LI K Q, GAO P Y, et al, 2022. Was dissolved nitrogen regime driving diatom to dinoflagellate shift in the Bohai Sea? Evidences from microcosm experiment and modeling reproduction[J]. Journal of Geophysical Research: Biogeosciences, 127(6): e2021JG006737. DOI:10.1029/2021JG006737 |
CICCHETTI D V, FEINSTEIN A R, 1990. High agreement but low kappa: Ⅱ. Resolving the paradoxes[J]. Journal of Clinical Epidemiology, 43(6): 551-558. DOI:10.1016/0895-4356(90)90159-M |
DAI A Q, LI K Q, DING D S, et al, 2015. Total maximum allocated load calculation of nitrogen pollutants by linking a 3D biogeochemical–hydrodynamic model with a programming model in Bohai Sea[J]. Continental Shelf Research, 111: 197-210. DOI:10.1016/j.csr.2015.08.028 |
DING X K, GUO X Y, ZHANG C, et al, 2020. Water conservancy project on the Yellow River modifies the seasonal variation of Chlorophyll-a in the Bohai Sea[J]. Chemosphere, 254: 126846. DOI:10.1016/j.chemosphere.2020.126846 |
EPPLEY R W, 1972. Temperature and phytoplankton growth in the sea[J]. Fishery Bulletin, 70(4): 1063-1085. |
FASHAM M J R, DUCKLOW H W, MCKELVIE S M, 1990. A nitrogen-based model of plankton dynamics in the oceanic mixed layer[J]. Journal of Marine Research, 48(3): 591-639. DOI:10.1357/002224090784984678 |
FEINSTEIN A R, CICCHETTI D V, 1990. High agreement but Low Kappa: I. The problems of two paradoxes[J]. Journal of Clinical Epidemiology, 43(6): 543-549. DOI:10.1016/0895-4356(90)90158-L |
GAO P Y, WANG P, CHEN S Q, et al, 2020. Effect of Ambient nitrogen on the growth of phytoplankton in the Bohai Sea: kinetics and parameters[J]. Journal of Geophysical Research: Biogeosciences, 125(12): e2020JG005643. DOI:10.1029/2020JG005643 |
JØRGENSEN S E, NIELSEN S N, JØRGENSEN L A, 1991. Handbook of Ecological Parameters and Ecotoxicology[M]. Amsterdam: Elsevier.
|
KRISTEK J, MOCZO P, GALIS M, 2010. Stable discontinuous staggered grid in the finite‐difference modelling of seismic motion[J]. Geophysical Journal International, 183(3): 1401-1407. DOI:10.1111/j.1365-246X.2010.04775.x |
LI K Q, MA Y P, DAI A Q, et al, 2017. Degradation dynamics and bioavailability of land-based dissolved organic nitrogen in the Bohai Sea: linking experiment with modeling[J]. Marine Pollution Bulletin, 124(2): 856-870. DOI:10.1016/j.marpolbul.2017.02.033 |
LI K Q, WANG X L, HAN X R, et al, 2009. Modelling nitrogen and phosphorus dynamics in a mesocosm pelagic ecosystem in Laizhou Bay in China[J]. Journal of Ocean University of China, 8(2): 133-140. DOI:10.1007/s11802-009-0133-6 |
LI X Y, YU R C, RICHARDSON A J, et al, 2023. Marked shifts of harmful algal blooms in the Bohai Sea linked with combined impacts of environmental changes[J]. Harmful Algae, 121: 102370. DOI:10.1016/j.hal.2022.102370 |
LIN G H, XU X F, WANG P, et al, 2020. Methodology for forecast and control of coastal harmful algal blooms by embedding a compound eutrophication index into the ecological risk index[J]. Science of the Total Environment, 735: 139404. DOI:10.1016/j.scitotenv.2020.139404 |
MOLL A, RADACH G, 2003. Review of three-dimensional ecological modelling related to the North Sea shelf system: Part 1: models and their results[J]. Progress in Oceanography, 57(2): 175-217. DOI:10.1016/S0079-6611(03)00067-3 |
NING X R, LIN C L, SU J L, et al, 2010. Long-term environmental changes and the responses of the ecosystems in the Bohai Sea during 1960-1996[J]. Deep Sea Research Part II: Topical Studies in Oceanography, 57(11/12): 1079-1091. |
RADACH G, MOLL A, 1993. Estimation of the variability of production by simulating annual cycles of phytoplankton in the central North Sea[J]. Progress in Oceanography, 31(4): 339-419. DOI:10.1016/0079-6611(93)90001-T |
REDFIELD A C, KETCHUM B H, RICHARDS F A, 1963. The influence of organisms on the composition of sea-water [M] // HILL M N. The Composition of Seawater: Comparative and Descriptive Oceanography. The Sea: Ideas and Observations on Progress in the Study of the Seas. New York: Interscience Publishers: 26-77.
|
RILEY G A, 1956. Oceanography of long island sound, 1952-1954. II. Physical oceanography[J]. Bulletin of the Bingham Oceanographic Collection, 15: 15-46. |
SAVCHUK O P, 2002. Nutrient biogeochemical cycles in the Gulf of Riga: scaling up field studies with a mathematical model[J]. Journal of Marine Systems, 32(4): 253-280. |
SKOGEN M D, MOLL A, 2000. Interannual variability of the North Sea primary production: comparison from two model studies[J]. Continental Shelf Research, 20(2): 129-151. |
WANG J J, YU Z G, WEI Q S, et al, 2019. Long-term nutrient variations in the Bohai Sea over the past 40 years[J]. Journal of Geophysical Research: Oceans, 124(1): 703-722. |
WEN Y H, PETERS R H, 1994. Empirical models of phosphorus and nitrogen excretion rates by zooplankton[J]. Limnology and Oceanography, 39(7): 1669-1679. |
XIN M, WANG B D, XIE L P, et al, 2019. Long-term changes in nutrient regimes and their ecological effects in the Bohai Sea, China[J]. Marine Pollution Bulletin, 146: 562-573. |