 2024, Vol. 55
2024, Vol. 55

浙江梅山湾表层沉积物中甲藻孢囊分布和多样性研究

http://dx.doi.org/10.11693/hyhz20230500115
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 冷天泽, 郭卓然, 邵倩文, 陆斗定, 戴鑫烽. 2024.
- LENG Tian-Ze, GUO Zhuo-Ran, SHAO Qian-Wen, LU Dou-Ding, DAI Xin-Feng. 2024.
- 浙江梅山湾表层沉积物中甲藻孢囊分布和多样性研究
- DISTRIBUTION AND DIVERSITY OF DINOFLAGELLATE CYSTS IN SURFACE SEDIMENTS IN MEISHAN BAY, ZHEJIANG
- 海洋与湖沼, 55(1): 145-154
- Oceanologia et Limnologia Sinica, 55(1): 145-154.
- http://dx.doi.org/10.11693/hyhz20230500115
文章历史
-
收稿日期:2023-05-31
收修改稿日期:2023-09-03
引用本文
冷天泽, 郭卓然, 邵倩文, 陆斗定, 戴鑫烽. 2024. 浙江梅山湾表层沉积物中甲藻孢囊分布和多样性研究[J]. 海洋与湖沼, 55(1): 145-154

LENG Tian-Ze, GUO Zhuo-Ran, SHAO Qian-Wen, LU Dou-Ding, DAI Xin-Feng. 2024. DISTRIBUTION AND DIVERSITY OF DINOFLAGELLATE CYSTS IN SURFACE SEDIMENTS IN MEISHAN BAY, ZHEJIANG[J]. Oceanologia et Limnologia Sinica, 55(1): 145-154.
浙江梅山湾表层沉积物中甲藻孢囊分布和多样性研究
1. 自然资源部第二海洋研究所 海洋生态系统动力学重点实验室 浙江杭州 310012;
2. 宁波海洋研究院 浙江宁波 315832
2. 宁波海洋研究院 浙江宁波 315832
摘要:甲藻孢囊可以为赤潮提供种源, 还可用于指示海区富营养化状态。以往对甲藻孢囊分布的研究多集中于开放性水域和自然形成的海湾中, 在半封闭性人工海湾中的研究较少。梅山湾原属南北开放水域, 但于2012~2017年在向陆侧建设了北坝和南坝, 使其成为半封闭式人工海湾。通过对梅山湾内外海域采集到的6份表层沉积物样品分析, 共鉴定出37种甲藻孢囊, 其丰度介于237~1 054 cysts/g。甲藻孢囊平均丰度湾内高于湾外, 推测是筑堤后湾内水动力减弱, 悬浮物质浓度降低, 水体透明度升高, 水中颗粒物质沉积速率降低和水体富营养化所导致。调查海域甲藻孢囊物种多样性指数介于1.63~2.47, 均匀度指数介于0.58~0.82, 两者湾外均显著高于湾内, 反映出湾内生态系统稳定性更弱, 发生赤潮的可能性更高。研究共检出16种赤潮种和9种有毒甲藻孢囊, 产毒种及赤潮种丰度和种类占比湾内都高于湾外, 优势种有原多甲藻(Protoperidinium sp.)、美利坚原多甲藻(Protoperidinium americanum)、透镜翼甲藻(Diplopsalis lenticula)、微小亚历山大藻(Alexandrium minutum)、链状裸甲藻(Gymnodinium catenatum)和锥状斯氏藻(Scrippsiella trochoidea)。“筑堤效应”加剧了湾内外表层沉积物中甲藻孢囊分布的差异性, 也加重了湾内有毒有害赤潮发生的风险, 故应重视对湾内有毒有害甲藻的监测。
关键词:有害赤潮 筑堤效应 半封闭人工海湾 甲藻孢囊
DISTRIBUTION AND DIVERSITY OF DINOFLAGELLATE CYSTS IN SURFACE SEDIMENTS IN MEISHAN BAY, ZHEJIANG
1. Second Institute of Oceanography, Ministry of Natural Resources, Key Laboratory of Marine Ecosystem Dynamics, Hangzhou 310012, China;
2. Ningbo institute of Oceanography, Ningbo 315832, China
2. Ningbo institute of Oceanography, Ningbo 315832, China
Abstract: Dinoflagellate cysts can serve as seed banks for algal blooms and be also used to indicate water eutrophication. Previous studies on the distribution of dinoflagellate cysts have predominantly focused on open waters and natural bays, with fewer investigations conducted in semi-enclosed artificial bays. Meishan Bay was once an open bay and accessible from the north and south, and underwent a transformation into a semi-enclosed artificial bay following the construction of dams from 2012 to 2017. In this study, six surface sediment samples were collected from the inner and outer waters of the bay. A total of 37 species of dinoflagellate cysts were identified in abundance ranging from 237 to 1 054 cysts/g. The average cyst abundance was higher in the inner water than in the outer water due to the decreases in suspended matter content, hydrodynamics, and sedimentation rate, and the increases in water transparency and the water eutrophication inside the bay. The Shannon-Wiener diversity index(H′) ranged between 1.63~2.47, and Pielou's evenness index(J′) ranged between 0.58~0.82. Both indices were significantly higher outside than inside of the bay, reflecting lower ecosystem stability and a higher possibility of red tides inside the bay. A total of 16 species capable of causing red tides and 8 toxic species of dinoflagellate cysts were identified in this study. Both the percentage of abundance and species number of toxic and red tide dinoflagellate cysts were higher inside the bay than outside. Dominant species included Protoperidinium sp., Protoperidinium americanum, Diplopsalis lenticula, Alexandrium minutum, Gymnodinium catenatum, and Scrippsiella trochoidea. The construction of dams not only exacerbated the disparity in dinoflagellate cysts in surface sediments between inner and outer regions of the bay but also increased the risk of toxic and harmful red tides within the bay. Therefore, monitoring toxic and harmful dinoflagellate within the bay shall be strengthened in the future.
Key words:
harmful algal blooms the effect of construction of dam semi-enclosed artificial bay dinoflagellate cyst
据统计, 75%左右的有毒有害赤潮由甲藻引起, 而在2 000多种有描述的海洋甲藻中超过10%的甲藻能产生孢囊(Head, 1996; Gómez, 2012)。甲藻孢囊具有较强的耐受不良环境能力, 在海底沉积环境中可存活数十年甚至上百年(Gómez, 2012; Ellegaard et al, 2018)。表层沉积物中的孢囊是过去一段时期内沉积的所有甲藻孢囊的集合(黄海燕等, 2009), 在遇到合适的环境条件后会萌发, 可为赤潮的发生提供“种源”(Anderson et al, 2014)。而不同年代沉积物中的甲藻孢囊可一定程度上反映该区域水体的富营养化历史(王朝晖, 2007a), 同时也为有毒有害赤潮发生历史和预测提供研判依据(Anderson et al, 2014)。
梅山湾作为宁波舟山港的重要组成部分之一, 与陆地联系紧密, 原先为南北开放的水道, 于2012~ 2017年期间在向陆侧建设了北坝和南坝, 合龙后形成了半封闭式的人工泻湖(孙丽梅等, 2018), 堤坝改变了湾内水动力环境, 使得湾内水体较为平静, 泥沙含量减少(余湾, 2018)。随着产业的升级, 梅山湾两侧陆地区域由渔业和农业向工业化发展, 形成了一个以石化、钢铁、能源、造纸和船舶等为重点的产业聚集带, 湾内则是以旅游、休闲和生态保护等为主的集聚区(王庆丽等, 2016)。建堤后, 梅山湾曾多次发生赤潮, 在2018年达到顶峰(Shao et al, 2020)。为探究建堤对该海湾生态系统的影响, 本研究在梅山湾内外海域共设置了6个调查站位, 对表层沉积物中的甲藻孢囊, 特别是有毒有害种开展了一系列研究, 同时对造成湾内外甲藻孢囊分布差异的原因进行了分析。
1 材料与方法 1.1 样品采集在梅山湾内外海域设置了6个采样点(图 1, 表 1), 于2022年11月下旬用彼得逊抓斗式采样器采集了表层沉积物样品。取上表层0~2 cm间的沉积物样品, 置于封口袋密封, 低温避光保存, 带回实验室后4 ℃冷藏至处理。
 |
| 图 1 梅山湾孢囊采样站位分布 Fig. 1 The sampling stations of cysts in Meishan Bay 注: 图中灰白区域始于2012年填海建造而成 |
表 1 梅山湾采样站位信息 Tab. 1 Information of the sampling stations in Meishan Bay
| 站位点 | 经度/E | 纬度/N | 水深/m | 位置 |
| S1 | 122°2′13.344″ | 29°49′43.032″ | 2.2 | 湾外 |
| S2 | 121°59′13.452″ | 29°48′51.624″ | 8.0 | 湾内 |
| S3 | 121°58′0.659″ | 29°48′0.216″ | 9.5 | 湾内 |
| S4 | 121°56′45.708″ | 29°46′55.993″ | 6.2 | 湾内 |
| S5 | 121°55′8.616″ | 29°45′21.744″ | 5.1 | 湾内 |
| S6 | 121°54′52.632″ | 29°44′4.74″ | 4.6 | 湾外 |
参考Bolch (1997)的方法进行样品处理, 依次经过“超声、过筛、离心和重液浮选”步骤。具体如下: 称取20 g左右沉积物样品, 混合均匀后分成两份。一份于称量瓶中称重, 经70 ℃烘箱中24 h后再次称重, 计算沉积物含水量; 另一份用于甲藻孢囊分析鉴定, 加入适量灭菌过滤海水稀释沉积物, 用超声波振荡2 min。依次通过125 µm和20 µm网筛, 用灭菌海水冲洗多次, 直至过滤液清澈。之后将20 µm网筛上的沉积物转移到20 mL玻璃管中, 然后利用胶头滴管多次向底部吹气, 使较重的沉积物颗粒首先沉降至试管底部, 用吸管吸取上浮液于10 mL离心管中, 重复多次以确保悬浮物都转移到离心管中, 最终定容至10 mL。之后通过重液浮选浓缩, 取15 mL离心管, 底部加入2 mL多聚钨酸钠(SPT)重液, 缓慢将定容后的悬浮液加入重液上方, 样品以3 000 r/min速率离心10 min, 去除部分上清液, 吸取交界面悬浮的甲藻孢囊于2 mL离心管中, 定容至2 mL, 保存于4 ℃冰箱中。根据样品清洁度, 吸取0.1~1 mL处理后的样品及适量过滤海水于计数框中, 使用倒置显微镜(Olympus CKX53, 日本)进行甲藻孢囊形态鉴定和数量统计, 使用正置显微镜(ZEISS Axiolab 5, 德国)拍摄照片。依据参考文献对孢囊进行定性定量分析(Matsuoka et al, 2000; 王朝晖, 2007b), 每个样品观察不少于100个甲藻孢囊, 甲藻孢囊丰度以每克干泥中的甲藻孢囊数量表示, 单位为cysts/g (干重)。
1.3 数据处理和统计分析孢囊的香农-威尔种类多样性指数(Shannon-Wiener diversity index, H′)(Shannon et al, 1949)、皮卢均匀度指数(Pielou’s evenness index, J′)(Pielou, 1969)计算公式为
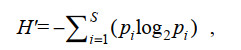 (1)
(1)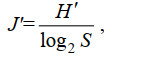 (2)
(2)优势种类的优势度指数(Y > 0.02)判断, 计算公式为
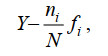 (3)
(3)式中, S为单一样品中的物种数, pi为第i种的细胞个体数与该样品的细胞个体总数的比值, ni为所有样品中第i种的细胞个体总数, N为所有样品的细胞个体总数, fi为第i种在所有样品中的出现频率。
使用IBM SPSS Statistics 23中的单因素方差分析(ANOVA)来分析湾内外不同类型物种在丰度和种数之间的差异性, 使用Surfer 15绘制采样站位、甲藻孢囊丰度、物种多样性指数和均匀度指数分布图, 使用Origin Pro 2021绘制柱状图。
2 结果与分析 2.1 甲藻孢囊种类组成和丰度分布在宁波梅山湾海域表层沉积物中, 共分析鉴定出甲藻孢囊37种, 其中多甲藻目孢囊18种、膝沟藻目10种、裸甲藻目7种和2种Thoracosphaerales目孢囊, 各站位所鉴定到的孢囊种类数为11~23种(表 2, 图 3)。甲藻孢囊丰度介于237~1 054 cysts/g, 站位平均甲藻孢囊丰度为606 cysts/g, S3站位甲藻孢囊丰度最高(1 054 cysts/g), S6站位最低(237 cysts/g) (图 2a)。各站位点平均丰度多甲藻目(368 cysts/g)高于膝沟藻目(106 cysts/g)和裸甲藻目(110 cysts/g)。物种多样性指数在1.63~2.47, S1站位最大, S3站位最小(图 2b); 均匀度指数在0.58~0.82, S6站位最大, S2站位最小(图 2c)。优势种有原多甲藻(Protoperidinium sp.)、美利坚原多甲藻(Protoperidinium americanum)、透镜翼甲藻(Diplopsalis lenticula)、微小亚历山大藻(Alexandrium minutum)、链状裸甲藻(Gymnodinium catenatum)和锥状斯氏藻(Scrippsiella trochoidea), 其中原多甲藻、美利坚原多甲藻、透镜翼甲藻和链状裸甲藻在所有站位均有分布(表 2)。根据参考文献, 37种甲藻孢囊中有19种为异养型, 16种为自养型, 其余2种孢囊营养类型不确定(Matsuoka, 1999; 王朝晖, 2007a; 石雅君等, 2011; Price et al, 2017; 李影等, 2017; Tian et al, 2018; 杨双, 2019; 钟文聪, 2019)。
表 2 梅山湾海域表层沉积物甲藻孢囊信息及分布状况 Tab. 2 Information and distribution of dinoflagellate cysts in surface sediments of the Meishan Bay
| 孢囊种类 | 站位 | |||||||||
| 目 | 中文名 | 藻类学名 | 孢粉学名 | 产毒 类型 |
S1 | S2 | S3 | S4 | S5 | S6 |
| 多甲藻目Peridiniales | ○原多甲藻 | Protoperidinium sp. | – | 37 | 19 | 52 | 50 | 101 | 36 | |
| ○原多甲藻 | Protoperidinium sp. | Brigantedinium irregulare | – | 11 | 2 | 0 | 0 | 0 | 0 | |
| ○原多甲藻 | Protoperidinium sp. | Trinovantedinium capitatum Reid | – | 6 | 0 | 2 | 0 | 0 | 0 | |
| ○美利坚原多甲藻 | Protoperidinium americanum | – | 53 | 34 | 44 | 13 | 16 | 13 | ||
| ○褐色原多甲藻 | Protoperidinium avellana | Brigantedinium cariacoense | – | 6 | 2 | 0 | 0 | 0 | 0 | |
| ○*窄角原多甲藻 | Protoperidinium claudicans | Votadinium spinosum | – | 6 | 0 | 0 | 0 | 0 | 0 | |
| ○*锥形原多甲藻 | Protoperidinium conicum | Selenopemphix quanta | – | 6 | 0 | 0 | 0 | 0 | 0 | |
| ○锥腹原多甲藻 | Protoperidinium conicoides | Brigantedinium simplex | – | 0 | 0 | 0 | 9 | 4 | 0 | |
| ○小齿原多甲藻 | Protoperidinium denticulatum | Brigantedinium sp. | – | 6 | 0 | 0 | 0 | 0 | 0 | |
| ○分支原多甲藻 | Protoperidinium divaricatum | Xandaradinium variabile | – | 6 | 0 | 8 | 0 | 4 | 7 | |
| ○*里昂原多甲藻 | Protoperidinium leonis | Quinquecuspis concreta | – | 0 | 3 | 4 | 7 | 0 | 0 | |
| ○点刺原多甲藻 | Protoperidinium punctulatum | Brigantedinium sp. | – | 6 | 0 | 0 | 0 | 0 | 0 | |
| ○*五边原多甲藻 | Protoperidinium pentagonum | Brigantedinium majusculum | – | 6 | 0 | 0 | 0 | 4 | 0 | |
| ○*赛裸原多甲藻 | Protoperidinium subinerme | Selenopemphix nephroides | – | 6 | 0 | 0 | 0 | 0 | 0 | |
| ○盾翼藻 | Zygabikodinium sp. | Dubridinium ulsterum | – | 0 | 0 | 0 | 5 | 0 | 0 | |
| ○微小盾翼藻 | Zygabikodinium lenticulatum | Dubridinium caperatum | – | 27 | 0 | 15 | 0 | 4 | 0 | |
| ○微小古生多甲藻 | Archaeperidinium minutum | – | 0 | 0 | 0 | 21 | 24 | 0 | ||
| ○透镜翼甲藻 | Diplopsalis lenticula | – | 95 | 381 | 453 | 284 | 235 | 71 | ||
| 膝沟藻目 Gonyaulacales |
▽安蒂奥条纹小锥藻 | Atlanticodinium striaticonulum | – | 22 | 5 | 0 | 0 | 0 | 10 | |
| ●巴哈马麦甲藻 | Pyrodinium bahamense var. bahamense | Polyspaeridium zoharyi | PSP | 0 | 0 | 8 | 5 | 0 | 4 | |
| ●易脆藻 | Fragilidium sp. | – | 0 | 0 | 0 | 13 | 0 | 0 | ||
| ●*多边舌甲藻 | Lingulodinium polyedrum | Lingulodinium machaerophorum | YTX | 0 | 5 | 0 | 0 | 0 | 4 | |
| ●*网状原角管藻 | Protoceratium reticulatum | Operculodinium centrocarpum | YTX | 0 | 2 | 0 | 0 | 0 | 0 | |
| ●膜状膝沟藻 | Gonyaulax membranacea | Spiniferites membranaceus | – | 0 | 0 | 2 | 0 | 0 | 0 | |
| ●*具指膝沟藻 | Gonyaulax digitale | Spiniferites bentori | – | 0 | 2 | 0 | 0 | 0 | 0 | |
| ●*具刺膝沟藻 | Gonyaulax spinifera | Spiniferites ramosus | YTX | 6 | 6 | 4 | 0 | 16 | 0 | |
| ●*塔玛/链状亚历山大藻 | Alexandrium tamarense/catenella | PSP | 0 | 22 | 8 | 5 | 4 | 7 | ||
| ●*微小亚历山大藻 | Alexandrium minutum | Alexandrium sp.2 (globe) | PSP | 0 | 52 | 303 | 75 | 16 | 29 | |
| 裸甲藻目Gymnodiniales | ○无纹多沟藻 | Polykrikos schwartzii | – | 6 | 0 | 0 | 0 | 8 | 0 | |
| ●*哈曼褐多沟藻 | Pheopolykrikos hartmannii | 鱼毒素 | 22 | 0 | 0 | 0 | 0 | 0 | ||
| ●*链状裸甲藻 | Gymnodinium catenatum | PSP | 164 | 91 | 99 | 29 | 43 | 49 | ||
| ●*伊姆裸甲藻 | Gymndinium impudicum | – | 0 | 12 | 44 | 13 | 4 | 0 | ||
| ●旋沟藻 | Cochlodinium sp.1 | – | 0 | 0 | 0 | 0 | 4 | 7 | ||
| ●旋沟藻 | Cochlodinium sp.2 | – | 6 | 0 | 0 | 0 | 0 | 0 | ||
| ▽*多环马格里夫藻 | Margalefidinium polykrikoides | – | 32 | 18 | 0 | 5 | 0 | 0 | ||
| Thoracosphaerales | ●斯氏藻 | Scrippsiella sp. | – | 6 | 3 | 0 | 0 | 0 | 0 | |
| ●*锥状斯氏藻 | Scrippsiella trochoidea | 有毒 | 48 | 35 | 8 | 9 | 31 | 0 | ||
| 孢囊种类数 | 23 | 18 | 15 | 15 | 16 | 11 | ||||
| 孢囊丰度/(cysts/g干重) | 589 | 694 | 1 054 | 543 | 518 | 237 | ||||
| 注: “●”表示自养; “○”表示异养; “▽”表示营养类型不确定; “*”表示赤潮种; PSP为麻痹性贝类毒素; YTX为虾夷扇贝毒素 | ||||||||||
 |
| 图 2 梅山湾各站位点甲藻孢囊相对丰度(a)、物种多样性指数(b)和均匀度指数(c)分布 Fig. 2 Relative abundance (a), Shannon-Wiener diversity index (H′) (b) and Pielou's evenness index (J′) (c) of dinoflagellate cysts in different stations of Meishan Bay |
 |
| 图 3 梅山湾海域表层沉积物中部分甲藻孢囊图集 Fig. 3 Photographs of partial dinoflagllate cysts in the surface sediments from Meishan Bay 注: a~b: 微小亚历山大藻; c: 塔玛(链状)亚历山大藻; d: 旋沟藻; e: 透镜翼甲藻; f: 具刺膝沟藻; g: 链状裸甲藻; h~i: 无纹多沟藻; j: 锥腹原多甲藻; k: 锥形原多甲藻; l: 里昂原多甲藻; m: 原多甲藻(孢粉学名Trinovantedinium capitatum); n: 网状原角管藻; o: 锥状斯氏藻; p: 微小盾翼藻。比例尺=10 µm |
每个站位都检出赤潮种类和有毒甲藻的孢囊(表 2), 有毒种包括产麻痹性贝类毒素的巴哈马麦甲藻(Pyrodinium bahamense var. bahamense)、塔玛(链状)亚历山大藻(Alexandrium tamarense/catenella)、微小亚历山大藻和链状裸甲藻, 产虾夷扇贝毒素的网状原角管藻(Protoceratium reticulatum)、多边舌甲藻(Lingulodinium polyedrum)和具刺膝沟藻(Gonyaulax spinifera), 产鱼毒素的哈曼褐多沟藻(Pheopolykrikos hartmannii)和对双壳类幼虫有致死效应的锥状斯氏藻(Tang et al., 2012)。
2.2 湾内湾外甲藻孢囊种类组成和丰度分布甲藻孢囊平均丰度湾内高于湾外(图 2a, 4a); 甲藻孢囊的物种多样性指数和均匀度指数湾内都显著低于湾外(图 4b, 4e); 孢囊平均种类数湾内外差异较小(图 4d); 有毒种平均丰度和种类占比湾内均高于湾外(图 4c, 4f); 赤潮种平均丰度和种类占比湾内均高于湾外(图 4c, 4f)。
 |
| 图 4 梅山湾内湾外甲藻孢囊平均丰度及种类组成(*: P < 0.05) Fig. 4 Average abundance and species composition of dinoflagellate cysts at the inner and outer stations of Meishan Bay (*: P < 0.05) |
本研究在梅山湾海域表层沉积物中鉴定出37种甲藻孢囊, 孢囊的种类在我国大多数海域检出数(15~50种)范围中偏高; 各站位点孢囊丰度介于237~1 054 cysts/g, 丰度范围与深圳盐田港相近, 比福建福宁湾, 福建东山湾, 南麂列岛和辽东半岛沿岸大, 比北黄海小; 多样性指数接近于福建东山湾和福建福宁湾, 低于深圳盐田港和胶州湾; 均匀度指数和福建东山湾、福建福宁湾和胶州湾相近, 略低于深圳盐田港(表 3)。
表 3 中国海域近岸表层沉积物中甲藻孢囊相关信息 Tab. 3 Relative informative of dinoflagellate cysts in surface sediments along Chinese coasts
| 采样时间 | 采样位置 | 孢囊种类数/种 | 自养/种 | 异养/种 | 孢囊丰度/(cysts/g) | 优势种 | 多样性指数 | 均匀度指数 | 来源 |
| 2022年12月 | 浙江梅山湾海域 | 37 | 16 | 19 | 237~1 054 | 原多甲藻、美利坚原多甲藻、透镜翼甲藻、微小亚历山大藻、链状裸甲藻和锥状斯氏藻 | 1.63~2.47 | 0.58~0.82 | 本研究 |
| 2020年四季 | 深圳 盐田港 |
38 | 异养型种类和丰度均高于自养型 | 297~996 | 锥状斯氏藻、亚历山大藻、链状裸甲藻、旋沟藻和多边舌甲藻为主导物种 | 均值3.65 | 均值0.93 | 赵薇等, 2022 | |
| 2018年4月 | 福建 福宁湾 |
25 | — | — | 178.6~386.3 | 别什藻 | 2.06~2.34 | 0.71~0.81 | 王朝晖等, 2022b |
| 2016年11月 | 福建 东山湾 |
22 | 14 | 8 | 70.6~136.4 | 原多甲藻 | 1.70~2.38 | 0.64~0.84 | 王朝晖等, 2022a |
| 2015年7月 | 胶州湾 | 39 | 20 | 17 | 膝沟藻 | 2.84~3.91 | 0.75~0.98 | 李影等, 2017 | |
| 2014年4、 6和8月 |
南麂 列岛 |
27 | 14 | 13 | 2~33 | 锥状斯氏藻 | 0.92~3.10 | — | 林顺利, 2016 |
| 2013年 | 辽东半岛沿岸 | 15 | 9 | 6 | 87~775 | — | — | — | 李大成等, 2019 |
| 2009年6月和2010年4~5月 | 北黄海 | 28 | 丰度中部海域多而近岸海域少 | 545~3 567 | 膝沟藻、塔马/链状亚历山大藻和微小亚历山大藻 | 1.77~3.31 | — | 石雅君等, 2011 | |
| 2007年4月和2008年 3月 |
大亚湾大鹏澳海域 | 50 | — | — | — | — | — | — | 付永虎等, 2011 |
梅山湾位于浙江省宁波市海域, 建堤前与东海直接相连, 长期受长江口、钱塘江和东海海潮的影响, 水体交换频繁, 泥沙含量较高(潘磊剑等, 2020; 闫慧贞等, 2021)。在修筑南北堤后, 梅山湾内悬浮泥沙浓度与建堤之前相比降低了约50~150 mg/L(余湾, 2018)。悬浮泥沙减少, 导致水体透明度增加, 水体光照得以增强, 这促进了各类浮游植物的生长, 如李振华等(2018)分析, 舟山海域春季浮游植物在近岸的丰度低于离岸丰度很可能是水体悬浮物浓度和浊度造成。另一方面, 两堤截流改变了梅山湾的潮汐和水动力环境(余湾, 2018), 使得湾内水动力减弱, 水体稳定性增加, 水中颗粒沉降速率降低。水体稳定性的增加可以提高浮游植物产量乃至藻华的发生(戴鑫烽等, 2014)。浮游植物产量的提升就会相应地提高甲藻孢囊的产量。同时, 梅山湾湾外水流复杂, 水体浑浊, 流速较快, 水中颗粒沉降速率较湾内高。研究表明, 水中颗粒沉降速率越高, 单位时间内形成的孢囊丰度越低(王朝晖等, 2022a), 如福建东山湾沉积物中孢囊丰度的差异主要和沉积物性质和沉积速率有关(王朝晖等, 2022a)。
水体富营养化程度也会影响表层沉积物中甲藻孢囊的丰度和群落结构。梅山湾水质富营养化程度湾内高于湾外(张美等, 2020), 已有结果表明, 富营养化程度较高的海区, 浮游植物的多样性指数和种类丰富度较低(李清雪等, 1999), 此外浮游植物生物多样性指数在赤潮区会显著低于非赤潮区(沈盎绿等, 2018)。本研究显示梅山海域湾内孢囊物种多样性指数和均匀度指数均显著低于湾外(图 3), 同时湾内曾多次暴发过赤潮(Shao et al, 2020), 反映出了湾内生态系统稳定性较弱。此外, 梅山湾海域表层沉积物中Hg的含量(0.73 mg/kg)高于象山港(0.11 mg/kg)、杭州湾(0.04 mg/kg)、浙江沿岸(0.04~0.05 mg/kg)、渤海湾(0.07 mg/kg)和辽东湾(0.07 mg/kg), 而Hg的主要来源是农业杀虫剂和污水(Wang et al, 2017; Jiang et al, 2018; Li et al, 2018; Zhao et al, 2018; Ding et al, 2019; Hao et al, 2019; Zhang et al, 2020), 这说明梅山海域水体环境受陆源影响较大。
影响表层沉积物中甲藻孢囊的环境因子众多, 除本文提到的水体透明度、水体动力环境和营养水平之外, 温度、盐度、人类活动、洋流变化和采样方式等因素也会影响甲藻孢囊群落组成(林顺利, 2016; 尤胜炮等, 2016)。
3.2 梅山湾有毒有害甲藻孢囊特征及潜在危害本研究共鉴定出16种赤潮种和9种有毒种甲藻的孢囊。包括产麻痹性贝类毒素(PSP)的链状裸甲藻、微小亚历山大藻、塔玛(链状)亚历山大藻和巴哈马麦甲藻的孢囊(Suzuki et al, 2003; Usup et al, 2012; Gu et al, 2013; Zou et al, 2014; Lewis et al, 2018), PSP毒素在贝类中毒反应中毒性最强, 危害最重, 在沿海地区赤潮发生时容易引发中毒事件(张曌等, 2023)。本研究中, 链状裸甲藻孢囊(平均丰度80 cysts/g)作为优势种在所有站位点均检出, 链状裸甲藻是我国常见的赤潮种之一(张文, 2009), 该藻赤潮暴发期间造成过严重的中毒事件和经济损失(陈火荣, 2018), 且在赤潮暴发后会形成孢囊, 为赤潮的再次发生提供种源(张文, 2009; 李光毅等, 2022)。微小亚历山大藻孢囊(平均丰度112 cysts/g)在所有湾内站位点均检出, 也是优势种之一。塔玛(链状)亚历山大藻是所有能够产生麻痹性贝类毒素的甲藻中分布最广泛、认知度最高的物种之一(Anderson et al, 2012), 能利用不同的氮源维持生长并持续暴发藻华(司冉冉等, 2017)。巴哈马麦甲藻孢囊在中国海域检出不多, 但野外调查发现其孢囊可为该赤潮暴发提供种源(Likumahua et al, 2021)。另外, 具有高麻痹性贝类毒素的有害赤潮常常发生在富营养化程度较高的水体环境中(汪宇等, 2021)。研究显示, 水体富营养化会促进链状裸甲藻的生长(胡蓉等, 2012), 而一定程度上的富营养化会使微小亚历山大藻胞内胞外毒素含量和毒性增加(刘丽君, 2020), 高氮磷比会促进微小亚历山大藻产毒效应(郝爽等, 2021)。在春季和秋季, 梅山湾湾外为中度污染, 主要为磷中等限制潜在性富营养以及贫营养, 湾内主要为轻度污染、磷限制潜在性富营养和磷限制富营养(张美等, 2020)。舟山海域的氮磷比长期严重失衡(李振华等, 2018), 这将提升浮游植物中的甲藻占比, 同时会使浮游植物细胞体积由大变小和产毒种类增加(Richardson, 1997; 宁修仁等, 2004)。因此很有必要加强对梅山湾海域尤其是湾内的监测, 以减少有毒有害赤潮造成的损失。
网状原角管藻、多边舌甲藻和具刺膝沟藻都是世界广布赤潮种, 这些藻极有可能是海底养殖扇贝中虾夷扇贝毒素的来源, 会导致扇贝死亡(Satake et al, 1997; Draisci et al, 1999; Tubaro et al, 2004; Rhodes et al, 2006; Riccard et al, 2009; 庞敏等, 2022), 其中具刺膝沟藻含虾夷扇贝毒素最高(庞敏等, 2022), 为预防上述藻类造成的危害, 应加强检测。
4 结论(1) 2022年冬季梅山湾海域表层沉积物中鉴定出4目37种甲藻孢囊, 丰度介于237~1 054 cysts/g, 物种多样性指数在1.63~2.47, 均匀度指数在0.58~0.82。
(2) 甲藻孢囊平均丰度湾内高于湾外, 物种多样性指数和均匀度指数湾内均显著低于湾外, 反映了湾内生态系统稳定性较弱。引起上述现象的原因可能是“筑堤效应”, 即筑堤降低了梅山湾的水动力, 导致湾内悬浮物质浓度降低, 水体透明度提高和水中颗粒沉降速率降低; 同时, 由于水交换减弱, 陆地污染源的排放加深了湾内水体富营养化程度, 为藻类繁殖(甚至是赤潮爆发)提供了充足营养。
(3) 本次调查共检出了16种赤潮藻和9种有毒甲藻孢囊, 有毒种及赤潮种甲藻孢囊丰度和种类占比湾内均高于湾外。其中, 产麻痹性贝类毒素的链状裸甲藻和微小亚历山大藻既是优势种, 也分布于所有湾内站位, 应该重视对湾内有毒有害甲藻的监测。
参考文献
王庆丽, 金旭孟, 方孟军, 2016. 梅山岛: 前世与今生[EB/OL]. (2016-11-29) [2023-04-14]. https://town.zjol.com.cn/system/2016/11/29/021384156.shtml.
|
王朝晖, 2007a. 中国典型近海海域甲藻孢囊分布及其与富营养化和赤潮生消关系研究[D]. 广州: 暨南大学.
|
王朝晖, 2007b. 中国沿海甲藻孢囊与赤潮研究[M]. 北京: 海洋出版社, 210.
|
王朝晖, 张宇宁, 王文婷, 等, 2022a. 福建东山湾表层沉积物中甲藻孢囊分布研究[J]. 热带海洋学报, 41(4): 154-162. |
王朝晖, 郑虎, 王文婷, 等, 2022b. 福建福宁湾表层沉积物中甲藻孢囊分布与多样性研究[J]. 海洋与湖沼, 53(6): 1396-1404. |
尤胜炮, 高寒, 雷向东, 等, 2016. 南麂列岛海域沉积物中甲藻孢囊的多样性和分布[J]. 海洋与湖沼, 47(2): 460-467. |
石雅君, 刘东艳, 邵红兵, 等, 2011. 北黄海表层沉积物中甲藻孢囊的分布特征[J]. 海洋通报, 30(3): 320-327. DOI:10.3969/j.issn.1001-6392.2011.03.014 |
付永虎, 王朝晖, 康伟, 等, 2011. 大亚湾大鹏澳海域表层沉积物中浮游植物休眠体集群的季节变化[J]. 台湾海峡, 30(1): 27-35. |
宁修仁, 史君贤, 蔡昱明, 等, 2004. 长江口和杭州湾海域生物生产力锋面及其生态学效应[J]. 海洋学报, 26(6): 96-106. DOI:10.3321/j.issn:0253-4193.2004.06.011 |
司冉冉, 关万春, 蔡景波, 等, 2017. 氮源对塔玛亚历山大藻生长和毒性的影响[J]. 生态学杂志, 36(10): 2880-2885. |
刘丽君, 2020. 基于液质联用分析的富营养化和酸化环境对微小亚历山大藻生长和产毒的影响[D]. 济南: 山东大学.
|
闫慧贞, 徐邹洋, 陈文杰, 等, 2021. 环梅山岛海域春季浮游古菌群落空间分布特征研究[J]. 微生物学报, 61(9): 2709-2725. |
孙丽梅, 汪文东, 高凤, 等, 2018. 宁波市某人工泻湖浮游生物变化特征及影响因素分析[J]. 生态毒理学报, 13(4): 60-67. |
李大成, 邵魁双, 巩宁, 等, 2019. 甲藻孢囊在辽东半岛沿岸海域表层沉积物中的分布[J]. 水产科学, 38(4): 545-549. |
李光毅, 刘保清, 邱勇, 等, 2022. 链状裸甲藻赤潮消亡过程及其与休眠包囊关系的初步研究[J]. 海洋学报, 44(8): 123-132. |
李振华, 周永东, 张亚洲, 等, 2018. 2014年春季舟山海域浮游植物群落特征[J]. 浙江海洋大学学报(自然科学版), 37(5): 405-411. |
李清雪, 陶建华, 1999. 应用浮游植物群落结构指数评价海域富营养化[J]. 中国环境科学, 19(6): 548-551. |
李影, 汤亚楠, 沈萍萍, 等, 2017. 胶州湾表层沉积物中甲藻孢囊的分布[J]. 海洋与湖沼, 48(4): 760-766. |
杨双, 2019. 渤海湾甲藻包囊研究[D]. 天津: 天津科技大学.
|
余湾, 2018. 基于遥感的梅山水道北堤对悬浮泥沙分布的影响研究[D]. 舟山: 浙江海洋大学.
|
汪宇, 张海燕, 隋延鸣, 等, 2021. 塔玛亚历山大藻产麻痹性贝类毒素能力的研究[J]. 海洋渔业, 43(3): 359-369. DOI:10.3969/j.issn.1004-2490.2021.03.011 |
沈盎绿, 欧阳珑玲, 尹艳娥, 等, 2018. 红色赤潮藻主导的混合赤潮对浮游生物群落结构的影响——以浙江南部沿海为例[J]. 海洋环境科学, 37(5): 625-630. |
张文, 2009. 不同环境因子对有害赤潮生物链状裸甲藻的生长和产毒的影响[D]. 广州: 暨南大学.
|
张美, 毛硕乾, 楼巧婷, 等, 2020. 2018年梅山湾营养盐的时空变化及富营养化分析[J]. 海洋环境科学, 39(6): 853-859. |
张曌, 栾玉静, 韩昱哲, 等, 2023. 麻痹性贝类毒素研究进展[J]. 刑事技术, 48(4): 418-425. |
陈火荣, 2018. 2017年福建海域链状裸甲藻赤潮事件应急处置与思考[J]. 渔业研究, 40(4): 308-314. |
林顺利, 2016. 春夏季南麂列岛海域甲藻孢囊分布特征与影响因素关系研究[D]. 舟山: 浙江海洋大学.
|
庞敏, 马新, 吴忠迅, 等, 2022. 海洋脂溶性藻毒素虾夷扇贝毒素产毒来源、毒性效应和检测技术研究进展[J]. 海洋湖沼通报, 44(6): 192-200. |
赵薇, 周进, 徐轶肖, 2022. 深圳盐田港海域甲藻孢囊的分布特征[J]. 应用与环境生物学报, 28(6): 1408-1414. |
郝爽, 刘丽君, 陈军辉, 等, 2021. 高氮磷比与酸化共同作用对微小亚历山大藻生长和产毒的影响[J]. 海洋科学, 45(2): 1-10. |
胡蓉, 徐艳红, 张文, 等, 2012. N、P、Mn和Fe对链状裸甲藻生长和产毒的影响[J]. 海洋环境科学, 31(2): 167-172. |
钟文聪, 2019. 渤海中部海域表层沉积物生源要素及浮游植物休眠体分布与分子多样性研究[D]. 广州: 暨南大学.
|
黄海燕, 陆斗定, 2009. 甲藻孢囊研究进展[J]. 海洋学研究, 27(3): 85-92. |
潘磊剑, 郭碧云, 2020. 近海表层水体悬浮泥沙遥感定量监测研究——以舟山海域为例[J]. 中国水运(下半月), 20(6): 224-226, 229. |
戴鑫烽, 陆斗定, 夏平, 等, 2014. 2010-2011年东海藻华高发区水体层化对东海原甲藻(Prorocentrum donghaiense Lu)藻华的影响[J]. 海洋与湖沼, 45(2): 217-224. |
ANDERSON D M, ALPERMANN T J, CEMBELLA A D, et al, 2012. The globally distributed genus Alexandrium: multifaceted roles in marine ecosystems and impacts on human health[J]. Harmful Algae, 14: 10-35. DOI:10.1016/j.hal.2011.10.012 |
ANDERSON D M, KEAFER B A, KLEINDINST J L, et al, 2014. Alexandrium fundyense cysts in the Gulf of Maine: long-term time series of abundance and distribution, and linkages to past and future blooms[J]. Deep Sea Research Part Ⅱ: Topical Studies in Oceanography, 103: 6-26. DOI:10.1016/j.dsr2.2013.10.002 |
BOLCH C J S, 1997. The use of sodium polytungstate for the separation and concentration of living dinoflagellate cysts from marine sediments[J]. Phycologia, 36(6): 472-478. DOI:10.2216/i0031-8884-36-6-472.1 |
DING X G, YE S Y, LAWS E D, et al, 2019. The concentration distribution and pollution assessment of heavy metals in surface sediments of the Bohai Bay, China[J]. Marine Pollution Bulletin, 149: 110497. DOI:10.1016/j.marpolbul.2019.110497 |
DRAISCI R, FERRETTI E, PALLESCHI L, et al, 1999. High levels of yessotoxin in mussels and presence of yessotoxin and homoyessotoxin in dinoflagellates of the Adriatic Sea[J]. Toxicon, 37(8): 1187-1193. DOI:10.1016/S0041-0101(98)00254-2 |
ELLEGAARD M, RIBEIRO S, 2018. The long-term persistence of phytoplankton resting stages in aquatic 'seed banks'[J]. Biological Reviews, 93(1): 166-183. DOI:10.1111/brv.12338 |
GÓMEZ F, 2012. A quantitative review of the lifestyle, habitat and trophic diversity of dinoflagellates (Dinoflagellata, Alveolata)[J]. Systematics and Biodiversity, 10(3): 267-275. DOI:10.1080/14772000.2012.721021 |
GU H F, LIU T T, VALE P, et al, 2013. Morphology, phylogeny and toxin profiles of Gymnodinium inusitatum sp. nov., Gymnodinium catenatum and Gymnodinium microreticulatum (Dinophyceae) from the Yellow Sea, China[J]. Harmful Algae, 28: 97-107. DOI:10.1016/j.hal.2013.06.001 |
HAO Z, CHEN L H, WANG C L, et al, 2019. Heavy metal distribution and bioaccumulation ability in marine organisms from coastal regions of Hainan and Zhoushan, China[J]. Chemosphere, 226: 340-350. DOI:10.1016/j.chemosphere.2019.03.132 |
HEAD M J, 1996. Modern dinoflagellate cysts and their biological affinities [M] // JANSONIUS J, MCGREGOR D C. Palynology: Principles and Applications. Dallas: American Association of Stratigraphic Palynologists Foundation: 1197-1248.
|
JIANG Q T, HE J Y, YE G Q, et al, 2018. Heavy metal contamination assessment of surface sediments of the East Zhejiang coastal area during 2012-2015[J]. Ecotoxicology and Environmental Safety, 163: 444-455. DOI:10.1016/j.ecoenv.2018.07.107 |
LEWIS A M, COATES L N, TURNER A D, et al, 2018. A review of the global distribution of Alexandrium minutum (Dinophyceae) and comments on ecology and associated paralytic shellfish toxin profiles, with a focus on Northern Europe[J]. Journal of Phycology, 54(5): 581-598. DOI:10.1111/jpy.12768 |
LI F P, MAO L C, JIA Y B, et al, 2018. Distribution and risk assessment of trace metals in sediments from Yangtze River estuary and Hangzhou Bay, China[J]. Environmental Science and Pollution Research, 25(1): 855-866. DOI:10.1007/s11356-017-0425-0 |
LIKUMAHUA S, SANGIORGI F, DE BOER M K, et al, 2021. Dinoflagellate cyst distribution in surface sediments of Ambon Bay (eastern Indonesia): Environmental conditions and harmful blooms[J]. Marine Pollution Bulletin, 166: 112269. DOI:10.1016/j.marpolbul.2021.112269 |
MATSUOKA K, 1999. Eutrophication process recorded in dinoflagellate cyst assemblages–a case of Yokohama Port, Tokyo Bay, Japan[J]. Science of the Total Environment, 231(1): 17-35. DOI:10.1016/S0048-9697(99)00087-X |
MATSUOKA K, FUKUYO Y, 2000. Technical guide for modern dinoflagellate cyst study [M]. Tokyo, Japan: WESTPAC– HAB, Japanese Society for The Promotion of Science.
|
PIELOU E C, 1969. An Introduction to Mathematical Ecology[M]. New York: Wiley–Interscience.
|
PRICE A M, COFFIN M R S, POSPELOVA V, et al, 2017. Effect of nutrient pollution on dinoflagellate cyst assemblages across estuaries of the NW Atlantic[J]. Marine Pollution Bulletin, 121(1/2): 339-351. |
RHODES L, MCNABB P, DE SALAS, et al, 2006. Yessotoxin production by Gonyaulax spinifera[J]. Harmful Algae, 5(2): 148-155. DOI:10.1016/j.hal.2005.06.008 |
RICCARDI M, GUERRINI F, RONCARATI F, et al, 2009. Gonyaulax spinifera from the Adriatic Sea: toxin production and phylogenetic analysis[J]. Harmful Algae, 8(2): 279-290. DOI:10.1016/j.hal.2008.06.008 |
RICHARDSON K, 1997. Harmful or exceptional phytoplankton blooms in the marine ecosystem[J]. Advances in Marine Biology, 31: 301-385. |
SATAKE M, MACKENZIE L, YASUMOTO T, 1997. Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin[J]. Natural Toxins, 5(4): 164-167. DOI:10.1002/19970504NT7 |
SHANNON C E, WEAVER W, 1949. The Mathematical Theory of Communication[M]. Urbana IL: University of Illinois Press.
|
SHAO Q W, LIN Z Z, ZHOU C X, et al, 2020. Succession of bacterioplankton communities over complete Gymnodinium-diatom bloom cycles[J]. Science of the Total Environment, 709: 135951. DOI:10.1016/j.scitotenv.2019.135951 |
SUZUKI T, ICHIMI K, OSHIMA Y, et al, 2003. Paralytic shellfish poisoning (PSP) toxin profiles and short-term detoxification kinetics in mussels Mytilus galloprovincialis fed with the toxic dinoflagellate Alexandrium tamarense[J]. Harmful Algae, 2(3): 201-206. DOI:10.1016/S1568-9883(03)00042-8 |
TANG Y Z, GOBLER C J, 2012. Lethal effects of Northwest Atlantic Ocean isolates of the dinoflagellate, Scrippsiella trochoidea, on Eastern oyster (Crassostrea virginica) and Northern quahog (Mercenaria mercenaria) larvae[J]. Marine Biology, 159(1): 199-210. DOI:10.1007/s00227-011-1800-x |
TIAN C, DOBLIN M A, DAFFORN K A, et al, 2018. Dinoflagellate cyst abundance is positively correlated to sediment organic carbon in Sydney Harbour and Botany Bay, NSW, Australia[J]. Environmental Science and Pollution Research, 25(6): 5808-5821. DOI:10.1007/s11356-017-0886-1 |
TUBARO A, SOSA S, ALTINIER G, et al, 2004. Short-term oral toxicity of homoyessotoxins, yessotoxin and okadaic acid in mice[J]. Toxicon, 43(4): 439-445. DOI:10.1016/j.toxicon.2004.02.015 |
USUP G, AHMAD A, MATSUOKA K, et al, 2012. Biology, ecology and bloom dynamics of the toxic marine dinoflagellate Pyrodinium bahamense[J]. Harmful Algae, 14: 301-312. |
WANG J, YE S Y, LAWS E A, et al, 2017. Surface sediment properties and heavy metal pollution assessment in the shallow sea wetland of the Liaodong Bay, China[J]. Marine Pollution Bulletin, 120(1/2): 347-354. |
ZOU C, YE R M, ZHENG J W, et al, 2014. Molecular phylogeny and PSP toxin profile of the Alexandrium tamarense species complex along the coast of China[J]. Marine Pollution Bulletin, 89(1/2): 209-219. |
ZHANG M, CHEN G, LUO Z T, et al, 2020. Spatial distribution, source identification, and risk assessment of heavy metals in seawater and sediments from Meishan Bay, Zhejiang coast, China[J]. Marine Pollution Bulletin, 156: 111217. |
ZHAO B F, WANG X M, JIN H B, et al, 2018. Spatiotemporal variation and potential risks of seven heavy metals in seawater, sediment, and seafood in Xiangshan Bay, China (2011-2016)[J]. Chemosphere, 212: 1163-1171. |