 2024, Vol. 55
2024, Vol. 55

2022年6月珠江流域洪水对下游溶解有机物浓度和组成的影响

http://dx.doi.org/10.11693/hyhz20230700144
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 傅芳敏, 姚爱林, 明玥, 王宪业, 高磊. 2024.
- FU Fang-Min, YAO Ai-Lin, MING Yue, WANG Xian-Ye, GAO Lei. 2024.
- 2022年6月珠江流域洪水对下游溶解有机物浓度和组成的影响
- IMPACT OF THE BASIN FLOOD IN JUNE 2022 ON THE CONCENTRATION AND COMPOSITION OF DISSOLVED ORGANIC MATTER IN THE LOWER ZHUJIANG (PEARL) RIVER
- 海洋与湖沼, 55(2): 397-408
- Oceanologia et Limnologia Sinica, 55(2): 397-408.
- http://dx.doi.org/10.11693/hyhz20230700144
文章历史
-
收稿日期:2023-07-10
收修改稿日期:2023-11-07
引用本文
傅芳敏, 姚爱林, 明玥, 王宪业, 高磊. 2024. 2022年6月珠江流域洪水对下游溶解有机物浓度和组成的影响[J]. 海洋与湖沼, 55(2): 397-408

FU Fang-Min, YAO Ai-Lin, MING Yue, WANG Xian-Ye, GAO Lei. 2024. IMPACT OF THE BASIN FLOOD IN JUNE 2022 ON THE CONCENTRATION AND COMPOSITION OF DISSOLVED ORGANIC MATTER IN THE LOWER ZHUJIANG (PEARL) RIVER[J]. Oceanologia et Limnologia Sinica, 55(2): 397-408.
2022年6月珠江流域洪水对下游溶解有机物浓度和组成的影响
1. 华东师范大学河口海岸学国家重点实验室 上海 200241;
2. 国家海洋环境监测中心国家环境保护海洋生态环境整治修复重点实验室 辽宁大连 116023
2. 国家海洋环境监测中心国家环境保护海洋生态环境整治修复重点实验室 辽宁大连 116023
摘要:当前极端气候事件频发, 引起了人们广泛的关注。然而, 气候变化对中国典型河流溶解有机物(DOM)的影响尚且未得到充分的认识。2021年11月至2022年10月, 每月于珠江下游广州段采集河水样品, 并分析其中溶解有机碳(DOC)、发色溶解有机物(CDOM)和荧光溶解有机物(FDOM)的浓度和组成。采样期间, 2022年6月珠江流域遭遇百年一遇的洪水。结果显示, 洪水大幅度降低了河水中的DOC和CDOM浓度, 并提高了DOM的芳香化程度。尽管洪水对DOM浓度产生了明显的稀释效应, 但通过分析FDOM组成, 进一步发现FDOM中不同组分对洪水的响应存在较大差异。FDOM短激发波长(230~235 nm)处的类蛋白质组分峰值在洪水期间出现高值; 与此同时, 长激发波长(280~285 nm)的类蛋白质组分和激发波长在345 nm处的类腐殖质组分峰值在洪水期间出现最低值。此外, 与长江下游相比, 珠江下游水体中往往具有较高的DOC和CDOM浓度、DOM芳香化程度以及CDOM分子量。研究将有助于进一步了解珠江等世界大河DOM浓度和组成的变化规律和控制机制, 以及揭示极端洪水对大河DOM动态变化产生的影响。
关键词:珠江 洪水 溶解有机碳 发色溶解有机物 荧光溶解有机物
IMPACT OF THE BASIN FLOOD IN JUNE 2022 ON THE CONCENTRATION AND COMPOSITION OF DISSOLVED ORGANIC MATTER IN THE LOWER ZHUJIANG (PEARL) RIVER
1. State Key Laboratory of Estuarine and Coastal Research, East China Normal University, Shanghai 200241, China;
2. State Environmental Protection Key Laboratory of Marine Ecosystem Restoration, National Marine Environmental Monitoring Center, Dalian 116023, China
2. State Environmental Protection Key Laboratory of Marine Ecosystem Restoration, National Marine Environmental Monitoring Center, Dalian 116023, China
Abstract: The recent frequent occurrence of extreme climate events has attracted widespread attention. However, the impacts of climate change on dissolved organic matter (DOM) in typical rivers in China are not yet well recognized. From November 2021 to October 2022, river water samples were collected monthly in Guangzhou section of the lower Zhujiang (Peral) River, and the concentration and composition of dissolved organic carbon (DOC), chromophoric dissolved organic matter (CDOM), and fluorescent dissolved organic matter (FDOM) were analyzed. During sampling period, the Zhujiang River basin encountered a "once in a century" flood in June 2022. Results show that the flood significantly reduced the concentrations of DOC and CDOM in the river water, and increased the DOM aromaticity. Although floods had a significant dilution effect on DOC and CDOM, further measurement of FDOM composition revealed that different FDOM components had different responses to the flood. The peaks of protein-like components at the shorter excitation wavelength (230~235 nm) reached its high value during the flood period. At the same time, the peaks of protein-like component with longer excitation wavelength (280~285 nm) and humic-like component (with excitation wavelength at 345 nm) showed the lowest values. In addition, compared with the lower Changjiang (Yangtze) River, the lower Zhujiang River showed generally greater concentrations of DOC and CDOM, and greater DOM aromaticity and molecular size. This study improved our understanding of the control mechanisms on the variations in DOM concentration and composition in the large rivers of the world like the Zhujiang River, and elucidated the impact of extreme flood on the DOM dynamics in large rivers.
Key words:
the Zhujiang (Pearl) River flood dissolved organic matter chromophoric dissolved organic matter fluorescent dissolved organic matter
河流是连接陆地和海洋的关键通道, 是陆源生源要素变化通过流域汇集后的集中体现区, 蕴含了丰富的气候变化和人类活动影响的信息, 在全球生物地球化学循环过程中扮演着重要的角色。溶解有机物(dissolved organic matter, DOM)和营养盐等生源要素在河流中的组成、来源、迁移转化途径及其受控因素一直是地球科学的重点关注问题(Guo et al, 2006; Worrall et al, 2007; Spencer et al, 2008; Seitzinger et al, 2010)。就径流量而言, 位于我国南部的珠江是仅次于长江的第二大河流, 其全长2 214 km, 流域面积45× 104 km2, 每年向南海输出300×109~370×109 m3的径流(年平均径流量为9 500~11 700 m3/s)和69×106~ 100×106 t的悬浮颗粒物(suspended particulate matter, SPM)(Callahan et al, 2004; Dai et al, 2008; Yang et al, 2011)。目前, 在人类活动、全球气候变化双重作用的影响和胁迫下, 我国长江、珠江、黄河等大河中生源要素的反馈机制及其时空变化规律始终是学术界相关领域的研究热点和难点(Zhang et al, 2007; 李奕洁等, 2015; Liu et al, 2020; Wu et al, 2021; Zhao et al, 2021; Liu et al, 2022; 王秀君等, 2022; Ming et al, 2023; Wang et al, 2023; Zhang et al, 2023)。
Zhao等(2021)和吴增斌等(2022)在长江下游的研究表明, 长江径流量可能是控制长江输出DOM浓度、通量及其组成的重要因素之一。Wang等(2022)的研究表明, 流域洪水通过改变长江向河口输出的SPM和营养盐通量, 进而影响长江口外海域赤潮事件发生的规模和频率。在珠江, 孙会国等(2006)和Tao等(2011)的研究也表明, 流域洪水事件可以极大地提高珠江颗粒有机碳(particulate organic carbon, POC)和溶解有机碳(dissolved organic carbon, DOC)的入海通量。由此可见, 极端洪水等气候事件通过改变径流量, 对长江、珠江等大河输出生源要素的浓度和组成有着调控作用。
2021年, 珠江流域汛期“当汛不汛”, 主要江河来水持续偏枯, 东江、韩江遭遇近60年来最严重干旱, 珠江三角洲各主要城市取水口受咸潮的影响加剧(钱燕等, 2022)。2021年冬至2022年春, 受有效降雨偏少、江河来水偏枯和水利工程蓄水不足等因素的影响, 珠江流域部分地区再次发生了区域性干旱(杜庆顺等, 2022)。而在随后的2022年5月下旬至7月上旬, 珠江流域则接连出现强降雨过程。在此期间珠江流域共发生两次流域性较大洪水, 特别是北江2022年第2号洪水, 形成时间与西江2022年第4号洪水接近, 导致两者产生叠加效应。此次系列洪水过程历时之长、量级之大历史罕见, 为1915年以来最大(陈学秋等, 2022; 钱燕等, 2023)。迄今为止, 该洪水对珠江输出的DOM等生源要素的浓度和组成所造成的影响及其程度, 尚且缺乏相关的研究报道。
与2022年6月类似, 在2005年6月下旬, 作为珠江最主要干流的西江就曾发生过一次重现期近200年的特大洪水事件。2005年6月10日, 高要站出现了24 000 m3/s的洪峰, 此后流量短时间消退, 但6月21日又开始急速上升, 并于6月24日达到最大洪峰(流量为54 900 m3/s), 6月28日流量迅速回落至25 000 m3/s以下(孙会国等, 2006)。孙会国等(2006)的研究还表明, 洪水期不到十日(2005年6月21日至28日), 西江径流、DOC和POC的通量就占到了全年各自通量的17%、25%和45%。Tao等(2011)的研究也表明, 2005年6月西江洪水期间输出的径流量占到了全年的36%, 输出的DOC占到了全年通量的30%。
本研究于2021年11月至2022年10月, 在珠江下游广州段固定站点进行了为期一年、每月一次的采样, 测定其中的DOC、发色溶解有机物(chromophoric dissolved organic matter, CDOM)、荧光溶解有机物(fluorescent dissolved organic matter, FDOM)等生源要素的相关参数。本研究旨在: (1) 了解DOM的浓度和组成在珠江下游广州河段的季节变化规律及其受控因素; (2) 揭示2022年6月珠江流域洪水对河流输出DOM的浓度和组成产生的影响; (3) 通过与长江的相关指标进行对比, 进一步了解珠江输出DOM的特征。
1 材料与方法 1.1 研究区域珠江由北江、东江、西江等三条主要支流构成, 其径流量分别占到了珠江总量的14%、8%和73% (Wei et al, 2014)。珠江下游入海口附近水系复杂, 河道纵横交错, 入海通道主要由八个口门构成(李博等, 2023)。由此, 珠江口具有“三江汇流、八口入海”的独特地貌特征, 是世界上最为复杂的河口之一(图 1)。珠江约50%~55%的径流通过东部的虎门、蕉门、洪奇门和横门输出到伶仃洋, 这四个口门输出径流分别约占伶仃洋接收水量的35%、33%、12%和20% (Callahan et al, 2004; He et al, 2010; Li et al, 2019), 而伶仃洋也在传统上被认为是珠江河口的主要区域(Chen et al, 2004b)。此外, 4月至9月被认为是珠江及其流域的洪季, 其径流量和泥沙通量大约分别占全年的70%~ 80%和95% (Chen et al, 2004a; Xia et al, 2004)。
 |
| 图 1 珠江流域和珠江口地图, 以及本研究采样点的位置 Fig. 1 Maps of the Zhujiang River Basin and the Zhujiang River estuary (left), and the location of the sampling site in this study region |
珠江上游地区喀斯特地貌广泛发育, 生态环境相对脆弱, 而下游珠江三角洲人口密集, 城市化程度高。珠江三角洲分布有广州、深圳、香港、澳门等大型城市(图 1)。近几十年来, 随着珠江三角洲城市化的快速发展, 人类活动对珠江流域和珠江口的影响与日俱增, 其对珠江向海输出陆源生源要素的通量和组成也产生了强烈的影响(Callahan et al, 2004; 张先毅等, 2020; Zhang et al, 2023)。据统计, 流域向珠江输出的废水约为9×109 m3/a, 其中包括城市废水7×109 m3/a和工业废水2×109 m3/a (Peng et al, 2017)。近几十年来, 基于人类活动和气候变化的影响(如20世纪90年代后珠江流域大量水库和大坝的修建), 珠江向海输出泥沙通量显著降低, 尤其是2000年以后, 龙滩水库的修建是西江输沙量进一步下降的最主要原因(Zhang et al, 2008; Wei et al, 2020; 杜以超等, 2022)。
虎门在八个口门中距离内陆最近(图 1), 受人类活动的影响最为直接, 是珠江(如东江、北江)流入外海的重要通道, 主要接纳广州市区和番禺区、佛山和东莞的大量居民生活污水和沿岸工农业废水的排放(曾丹娜等, 2020)。通过流经广州的虎门通道输出的大量废水对珠江口(伶仃洋)生态环境的影响尤其明显。根据逄勇等(2001), 虎门输出的有机污染物约占东部四个口门输出总量的70%。He等(2010)则进一步指出, 人类活动排放来源已经成为珠江DOC的重要组成部分; 在珠江广州段, 人类活动来源已经占到了DOC通量的54% (其余31%和15%分别为陆地有机物来源和水体浮游植物生产来源)。由此, 为了更聚焦于人类活动的影响, 本研究选择珠江虎门入海口上游的广州段作为采样站点。
1.2 样品采集于2021年11月至2022年10月期间, 在珠江广州段固定站点(经纬度为113°18′E和23°06′N, 图 1)进行每月一次的水样采集, 其中2022年4月与5月样品因新冠疫情未能采集, 2022年6月样品(采样时间: 6月25日)在洪水期内(5月30日至7月5日)采集。洪水期间, 西江第3号洪水与北江第1号洪水时间接近(分别为6月12日和6月14日), 促成6月16日珠江达到洪峰(流量为5.67×104 m3/s)(珠江径流量数据为北江石角站、东江博罗站和西江高要站日径流量之和, 数据来源于珠江水利委员会官网http://www.pearlwater.gov.cn/)。随后, 6月21日北江发生特大洪水, 样品于洪水发生后第四天采集, 受洪水充分影响(图 2)。在采样过程中, 用采样桶采集表层水样, 用1 L高密度聚乙烯瓶将样品避光保存, 带回实验室并于采样后12 h内完成过滤处理。在实验室, 使用预先高温灼烧(450 ℃和5 h)并称重的玻璃纤维膜(Whatman GF/F, 孔径为0.7 µm)过滤水样。其中, 用于测定SPM浓度的滤膜和用于测定DOC浓度的滤液于–20 ℃下冷冻保存直至测定。用于测定CDOM和FDOM的滤液于4 ℃下避光冷藏保存, 并在48 h内完成测定。实验涉及的采样桶、样品瓶和滤器均在使用前经pH < 2的稀盐酸溶液浸泡24 h以上, 并用Milli-Q水彻底冲洗。
 |
| 图 2 2022年4月1日至8月31日(洪水前后)珠江下游日径流量变化 Fig. 2 Variation in daily discharge in the lower Zhujiang River from April 1 to August 31, 2022 (before and after flood) |
样品盐度采用HACH盐度计(型号: HQ-40d)进行测定, 本研究涉及的10个样品盐度变化范围为0.09~ 1.96, 平均值为0.43±0.56。样品过滤后, 将过滤膜烘干(50 ℃和48 h)后测定其质量, 并将该值减去过滤前的空白膜质量, 该质量差值除以实际过滤体积得到样品的SPM质量浓度(单位为mg/L)。
样品的DOC浓度(单位为µmol/L)使用总有机碳分析仪器(日本岛津公司, 型号: TOC-VCPH)并依据高温燃烧法(680 ℃)进行测定。在分析前, 用高纯度盐酸对样品进行酸化, 使其pH < 2。每个测定日都建立新的标准曲线。为了保证样品质量和实时监控仪器状态, 样品测定过程中, 使用国际标准DOC深海样品(购自美国迈阿密大学)和Milli-Q水进行质量控制。
使用紫外-可见光分光光度计(北京普析通用公司, 型号: TU1810DPC)测定CDOM吸收光谱。测定的波长范围为200~800 nm, 波长间隔为1 nm。测定过程中使用光程为5 cm的石英比色皿。在本研究中, 使用254 nm处的光吸收系数a254 [单位为m−1, 见公式(1)]定量CDOM浓度。以往文献中, a254已经被广泛用于定量自然水体中的CDOM浓度(Zhang et al, 2018, 2021; Johnston et al, 2020; Yan et al, 2021; Zhou et al, 2021), 与300、350 nm等较长波长相比, 254 nm处的样品光吸收系数对于CDOM浓度变化的响应更灵敏。使用254 nm处单位距离(1 m)的吸光度(A254, 无量纲)与DOC浓度(此处使用的单位为碳的质量浓度, 即mg C/L)的比值SUVA254 [单位为L/(mg C∙m)]来表征DOM的芳香化程度(Weishaar et al, 2003)。此外, 使用S275~295 (即275~295 nm处的光吸收系数先取自然对数再进行线性拟合后直线的斜率, 单位为nm−1)和SR(即275~295 nm处S275~295与350~400 nm处S350~400两个数值的比值)表征CDOM的分子量, 一般认为S275~295和SR越高, CDOM分子量越小(Helms et al, 2008)。
计算各波长处光吸收系数a依据公式(1):
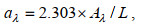 (1)
(1)其中λ为特定波长(单位为nm); A为特定波长处的吸光度(无量纲), 为了消除折射效应的影响, 该值通过减去Milli-Q水测定的650~800 nm处吸光度平均值进行修正; L为光程(单位为m)。
FDOM的荧光激发-发射三维光谱(excitation-emission matrix, EEM)使用荧光分光光度计(日本日立公司, 型号: F-2500)测定。测定过程中使用的石英比色皿的规格为1 cm × 1 cm。测定过程中, 激发波长的扫描范围为220~450 nm, 步长为5 nm。每一个激发波长下发射波长的扫描范围为230~600 nm, 测定时每2 nm进行一次读数。激发光和发射光的狭缝宽度均设定为5 nm。在测定FDOM时, 需考虑内滤效应(inner filter effect, IFE)导致的误差。根据Kothawala等(2013)的研究, 大多数自然水体样品无需稀释。因此, 通常采用样品的吸光度光谱计算内滤效应的校正因子I矩阵, 将经光谱校正的EEM与I矩阵对应相乘从而实现内滤效应校正(Murphy et al, 2010, 2013; Kothawala et al, 2013), 校正因子I的计算公式见公式(2):
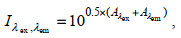 (2)
(2)其中, λex和λem分别代表激发波长和发射波长, Aλex和Aλem则分别表示激发波长和发射波长处的吸光度。每个测定日均使用当天新配制的0~6.4×10−2 µmol/L二水硫酸奎宁[(C20H24N2O2)2·H2SO4·2H2O]标准曲线, 测定的EEM每一位置的荧光读数均通过除以Ex/Em = 350/450 (nm/nm)处1.3×10−3 µmol/L浓度的二水硫酸奎宁的荧光值(即标准曲线的斜率)予以校正。本文报告的荧光值单位为µmol/L。
2 结果 2.1 DOC和CDOM全年观测期间, 珠江月平均径流量的变动范围为0.35×104~4.05×104 m3/s。径流量的最低值和最高值分别出现在2022年10月和2022年6月, 后者是前者的11.6倍。SPM浓度范围为9.9~67.6 mg/L(表 1), 最高值仍出现在径流量最大的6月, 最低值出现在径流量较低的3月, 最高值是最低值的6.8倍。受洪水期间SPM浓度较高的影响, 全年10个样品SPM浓度与月平均径流量之间呈显著正相关关系(P < 0.05)。
表 1 2021年11月至2022年10月珠江下游10个样品相关指标的测定结果 Tab. 1 Results of all parameters for the 10 samples collected in the lower Zhujiang River from November 2021 to October 2022
| 采样时间 | SPM/(mg/L) | DOC/(µmol/L) | a254/m–1 | SUVA254/[L/(mg C·m)] | S275~295/nm–1 | SR |
| 2021年11月28日 | 27.5 | 156 | 14.3 | 3.3 | 0.015 1 | 0.95 |
| 2021年12月18日 | 44.1 | 234 | 14.4 | 2.2 | 0.014 9 | 0.81 |
| 2022年1月9日 | 46.7 | 246 | 15.1 | 2.2 | 0.013 8 | 0.95 |
| 2022年2月24日 | 21.4 | 232 | 15.6 | 2.4 | 0.015 4 | 1.24 |
| 2022年3月27日 | 9.9 | 256 | 15.3 | 2.2 | 0.016 3 | 0.74 |
| 2022年6月25日 | 67.6 | 106 | 11.6 | 4.0 | 0.013 5 | 1.27 |
| 2022年7月26日 | 32.6 | 161 | 11.2 | 2.5 | 0.015 8 | 0.74 |
| 2022年8月24日 | 14.1 | 219 | 15.8 | 2.6 | 0.015 8 | 0.80 |
| 2022年9月20日 | 22.1 | 206 | 14.3 | 2.5 | 0.016 8 | 0.93 |
| 2022年10月29日 | 24.8 | 210 | 13.1 | 2.2 | 0.016 5 | 0.92 |
| 注: SPM为悬浮颗粒物浓度; DOC可用于表征DOM的相对浓度; a254表征CDOM的相对浓度; SUVA254表征DOM的芳香化程度; S275~295为DOM在275~295 nm波段的吸收光谱斜率; SR为S275~295与S350~400的比值 | ||||||
与月平均径流量和SPM浓度类似, DOC浓度也表现出了强烈的季节变化(表 1, 图 3)。2022年6月洪水发生期间, DOC浓度远远低于其他月份, 仅为106 µmol/L, 其次为11月和7月(洪水后期), 浓度分别为156和161 µmol/L, 而其余七个样品浓度均在206 µmol/L以上。2022年3月DOC浓度呈现最高值为256 µmol/L, 该值为洪水期间(2022年6月) DOC浓度的2.4倍。
 |
| 图 3 2021年11月至2022年10月珠江下游月平均径流量、DOC浓度以及a254值的变化 Fig. 3 Variations in monthly average discharge, DOC concentration, and a254 values in the lower Zhujiang River from November 2021 to October 2022 |
表征CDOM浓度的a254在洪水期间(2022年6月)和洪水后期(2022年7月)也出现了远低于其他月份的值, 这一点与DOC浓度类似(表 1和图 3)。洪水后期a254值(11.2 m–1)甚至略低于洪水期(11.6 m–1), 前者也是所有10个样品中a254的最低值。8月出现全年最高值(15.8 m–1), 仅为最低值的1.4倍。上述比值说明, 相较于径流量、SPM浓度和DOC浓度等指标, CDOM浓度在全年的季节变化幅度最小。
SUVA254的变化范围为2.2~4.0 L/(mg C∙m)(表 1), 其中最大值出现在洪水期(6月), 最小值出现在12、1、3和10月。6月最高值的出现说明珠江洪水期间河水DOM具有较高的芳香性(Weishaar et al, 2003)。全年S275~295的变化范围为0.013 5~0.016 8 nm–1 (表 1), 最低值出现在1和6月, 而最高值出现在9和10月。SR全年的变化范围为0.74~1.27 (表 1), 最高值出现在6月, 而最低值则出现在3月以及洪水后期的7月。S275~295和SR这两个参数都可以用来指示CDOM的分子量大小, 两个参数值越高, 对应的CDOM分子量往往越小(Helms et al, 2008)。洪水期间S275~295的低值表明洪水输出的较高芳香性的DOM(或CDOM)具有较大的分子量; 而SR的结果则相反, 最高值同样出现在6月, 表明该月份CDOM分子量最低(Helms et al, 2008)。需要指出的是, Helms等(2008)的研究也表明这两个参数的变化确实还受到了除分子量以外其他因素的显著影响。后续需要进一步开展研究来明确S275~295和SR对CDOM分子量的指示意义和各自的局限性, 以及其他可能影响这两个参数变化的因素。
2.2 FDOM10个样品的荧光EEM测定结果显示, 珠江下游广州段采样站点全年FDOM主要由三个峰(组分)构成(图 4)。第一个组分(称为C1)激发和发射光位置主要集中在Ex/Em=(230~235 nm)/(340~354 nm)处, 第二个组分(称为C2)的位置主要集中在Ex/Em=(280~285 nm) /(330~354 nm)处。与此同时, 在更长波长范围内还有一个荧光强度较低的第三个组分(称为C3), 位置在Ex/Em = (345 nm)/(416~426 nm)附近。其中, C1峰在洪水期间6月的样品中占主导的地位, 峰值为全年第三高值; 而C3峰在6月的样品中最不明显(图 4)。此外, 在11月和6月至8月的四个样品中, C1峰明显高于C2峰(C1/C2峰高比值在2.06~4.37的范围内); 而在其他六个样品中, 尽管C1峰仍高于C2峰, 但两者峰高差距相对较小(C1/C2峰高比值在1.60~1.95的范围内)(图 4)。
 |
| 图 4 2021年11月至2022年10月珠江下游采样站点采集的10个样品的荧光激发-发射三维光谱 Fig. 4 Fluorescent excitation-emission spectra of the 10 samples collected in the lower Zhujiang River from November 2021 to October 2022 |
根据Coble (1996)对EEM中各处荧光峰位置的研究和命名, 本研究的C2峰是以类色氨酸为主的类蛋白质T峰, 本研究的C3峰为类腐殖酸M峰。但由于Coble (1996)中FDOM测定的最低激发波长为260 nm, 因此其并未指明Ex/Em=(230~235 nm)/(340~ 354 nm)处(即本研究的C1峰)指示的为何种DOM物质。Mann等(2012)中EEM测定最低激发波长设定为240 nm, 指出该处荧光峰是与T峰(即本研究的C2峰)一同出现的类蛋白质物质。Coble (2013)通过延展激发波长测定下限进一步明确了该位置的荧光峰确实指示的是类蛋白质DOM物质, 并将其命名为AT峰。
尽管上述研究认为C1和C2峰均可代表珠江输出DOM中的类蛋白质组分, 但本研究全年10个样品的结果表明, 洪水对C1和C2两个组分的影响存在明显差异。其中C1组分峰值全年的变动范围为152× 10–3~263×10−3 µmol/L, 洪水期间(6月)出现C1峰的高值。C2和C3峰值在全年的变动范围分别为57×10–3~159×10−3 µmol/L和26×10–3~57×10−3 µmol/L, 两者的最低值也同样出现在洪水期间(图 4)。
3 讨论 3.1 洪水的影响张连凯等(2013)的研究表明, 珠江SPM、POC和DOC浓度在洪季升高的趋势明显: 以DOC为例, 与2012年4月相比, 洪季2012年7月东江博罗站、北江石角站、西江高要站以及西江上游的DOC浓度分别提高了21%、8%、24%和28%, 表明径流量的增加对DOC浓度的增长有促进作用(即冲刷效应)。Zhao等(2021)在研究长江下游2018年7月至2019年6月间DOC浓度的季节变化时, 也发现径流量对DOC浓度的冲刷效应; 但对于表征CDOM浓度的a254, Zhao等(2021)既没有观测到径流量的冲刷效应, 也没有观测到稀释效应。
图 5表明, 与其他月份相比, 6月珠江突增的径流量使得SPM浓度明显升高, 并在洪水后期快速下降。本研究DOC和CDOM浓度对于径流量的响应与张连凯等(2013)的研究结果不同, 洪水期间径流量对这两个指标表现为稀释作用, DOC和CDOM浓度明显低于其他月份, 并且直到洪水后期的7月, a254仍维持在接近于6月的低值水平上。2022年6月的洪水不仅改变了DOC和CDOM的浓度, 同时改变了它们的组成。SUVA254的结果进一步表明, 洪水期间带来的DOM分子具有更高的芳香化程度。对于FDOM, 洪水带来了更多的C1组分以及更少的C2和C3组分。
 |
| 图 5 珠江下游洪水期间(2022年6月)、洪水后(2022年7月)以及其他月份(取平均值)相关指标测定结果的比较 Fig. 5 Comparison of relevant indicators among samples collected during the flood (June 2022), after the flood (July 2022), and those of other months (averaged) |
在珠江口的研究中, Hong等(2005)在2002年11月采集了贯穿虎门淡水端元到珠江口(伶仃洋)的样品(该研究FDOM测定最低激发波长为210 nm), 也在虎门河道表层河水样品测得中发现, 最主要的峰出现在位置Ex/Em=(230 nm)/(348 nm)处(对应本文的C1组分), 并且他们发现该峰为所有样品(涵盖了从零盐度河水样品到高盐度珠江口海水样品)EEM中最主要的峰。上述结果与本研究全年在珠江下游广州段采集的样品的测定结果一致(图 4)。Hong等(2005)认为该类蛋白质组分主要来源于人类活动产生的废水, 反映了珠江乃至珠江口海域中DOM深受人类活动影响的情况。而在本研究的10个样品中, 除2021年12月、2022年1月外, 洪水时期(2022年6月)的C1峰值明显高于其他月份(图 4)。此次洪水主要由上游站位和本地的降雨共同导致, 因此C1峰值的升高可能受到洪水期间冲刷效应和污水溢流叠加影响, 从而导致洪水携带了更高比例和通量的通过人类活动产生的DOM进入珠江干流继而被输送到珠江口海域。
将FDOM的三个组分与珠江下游每月的平均径流量做相关性分析, 发现三个组分表现出较大差异。与DOC和CDOM浓度类似, C2和C3组分的最低值均出现在了洪水期间的2022年6月, 次低值出现在洪水后期的2022年7月。C2和C3峰值与月平均径流量之间呈现出显著的负相关关系(P < 0.05; P < 0.05) (图 6), 说明径流量对FDOM中C2和C3组分也主要表现出了稀释效应, 这一点与DOC和CDOM的趋势相一致。与C2和C3不同, 对于C1组分而言, 其高值出现在全年径流量最高的6月(图 6)。在洪水期间(2022年6月)径流量的升高对C1组分表现出了冲刷效应。然而, C1峰的荧光强度与月平均径流量之间没有显著的相关关系, 说明C1峰值可能受到其他因素影响。值得注意的是, C1峰作为FDOM三个峰中最主要、峰值最高的峰, 在洪水期间其荧光强度的增加并没有体现在DOC和CDOM浓度的增加上, 可能进一步说明不同的DOM定量指标分别侧重指示不同的DOM组分。
 |
| 图 6 2021年11月至2022年10月珠江下游采集的10个样品FDOM中的C1、C2、C3三个组分峰值与珠江下游月平均径流量的相关关系 Fig. 6 Correlations between the monthly mean discharge of the lower Zhujiang River and the peak values of C1, C2 and C3 components in FDOM (fluorescent dissolved organic matter) for the 10 samples collected in the lower Zhujiang River from November 2021 to October 2022 |
单森等(2020)分别于2016年3月至7月和2018年4月对长江干流17个站点和珠江干流4个站点进行了DOC样品的采集, 并测定了其中的δ13C和Δ14C值。单森等(2020)的结果表明, 两条河流DOC中δ13C值均在−28‰~−25‰; 对于DOC中的Δ14C值, 除了珠江最下游一个站点Δ14C约为−300‰外, 其余珠江和长江样品的Δ14C值均在−200‰~−100‰的范围内。该报道从DOC中δ13C和Δ14C值的角度对比了长江和珠江DOM组成的不同。
在本节中, 我们将珠江下游实测的数据与Zhao等(2021)研究中长江下游为期一年(2018年7月至2019年6月)的结果进行对比(图 7)。与长江相比, 珠江具有较低的SPM浓度、较高的DOC和CDOM浓度以及DOM芳香化程度(图 7)。此外, 图 7中S275~295和SR的结果均表明, 珠江DOM具有较大的分子量。Zhao等(2021)在对比世界范围内众多河流的DOC浓度、SUVA254、S275~295、SR等参数的关系发现, DOC浓度与DOM的芳香化程度越高时, 其分子量越大。本研究珠江和长江中DOM相关参数的比较结果均符合上述规律。
 |
| 图 7 珠江下游全年每月采集样品(2021年11月至2022年10月)与长江下游全年每月采集样品(2018年7月至2019年6月)相关指标平均值的对比 Fig. 7 Comparison in annual average values of relevant indicators between the monthly collected samples from the lower Zhujiang River (from November 2021 to October 2022) and those from the lower Changjiang River (from July 2018 to June 2019) |
在2020年7月, 长江流域也发生了百年一遇的洪水事件, 2020年7月长江大通站月平均径流量为71 300 m3/s, 为1960年以来仅次于长江大坝修建前的1998年8月(77 100 m3/s)和1999年7月径流量(73900 m3/s)的第三高值(数据来源于长江水文网http://www.cjh.com.cn/)。根据张淑坤等(2022)的研究, 在长江洪水期间(2020年7月13日至27日, 大通站日平均径流量为71 800~81 600 m3/s), 长江下游徐六泾站点的DOC浓度从 > 180 µmol/L (平常洪季)被稀释至最低值153 µmol/L (7月15日), 该浓度仍高于长江一般枯季时的浓度(140~150 µmol/L, 见Zhao et al, 2021)。与长江不同的是, 尽管珠江下游DOC浓度普遍高于长江下游(图 7), 但发生在2022年6月珠江的特大洪水将DOC浓度稀释至106 µmol/L (表 1, 图 3), 该值不仅低于珠江一般枯季的DOC浓度值, 甚至低于长江一般枯季出现的DOC季节性低值。由此可见: (1) 无论是长江还是珠江, 尽管一般洪季径流量的升高可能会导致DOC浓度的升高(冲刷效应), 但在极端洪水条件下径流量的进一步升高反而会大幅降低河水中DOC浓度, 产生稀释效应; (2) 根据两条大河DOC浓度的年内变化幅度, 特大洪水对珠江DOC浓度的影响可能要大于对长江DOC浓度的影响。
表征CDOM浓度的a254在长江下游徐六泾站全年的变化范围约为8~13 m–1 (Zhao et al, 2021)。由图 7可知, 珠江全年的a254值一般高于长江。非洪水期间珠江a254值一般高于13 m–1, 洪水通过稀释效应, 使得珠江下游a254值在洪水期间下降至11~12 m–1 (表 1, 图 3)。值得注意的是, 即使在洪水期间珠江的a254值仍高于长江下游a254的年内低值, 这一点和DOC浓度在两条大河间的变化有所不同。综上所述, 无论对于DOC还是CDOM, 其浓度和组成在长江和珠江之间表现出了明显的差异。此外, 流域特大洪水对长江和珠江DOM浓度和组成都产生了重要影响, 甚至可以决定相关参数在两条大河全年变化范围的上限和下限。
4 结论通过对珠江下游广州段固定站点2021年11月至2022年10月为期一年的每月采样, 本研究发现:
(1) 发生在2022年6月的特大洪水极大地降低了珠江河水中DOC和CDOM的浓度, 产生了明显的稀释效应。除了浓度变化外, 洪水显著提高了DOM的芳香化程度。
(2) 尽管总体上洪水对珠江DOM中大部分组分产生了稀释效应, 但DOM中不同组分的变化在洪水期间差异仍较大。2022年6月洪水期间, 短激发波长处的类蛋白质组分浓度不降反升, 径流量突增对该组分产生的冲刷效应超过了稀释效应。
(3) 与长江下游相比, 珠江下游河水中DOC和CDOM浓度、DOM芳香化程度和分子量相对较高。特大洪水对长江和珠江DOM浓度和组成的年内变化范围都表现出了重要的影响。
参考文献
王秀君, 王晓雯, 尤再进, 等, 2022. 近20年台风事件对黄渤海颗粒有机碳季节性的影响[J]. 海洋与湖沼, 53(4): 861-871. |
孙会国, 汉景泰, 张淑荣, 等, 2006. "05-06"西江特大洪水对河流碳输出通量的影响[J]. 科学通报, 51(23): 2773-2779. |
杜以超, 罗孝文, 王峻, 等, 2022. 近70年珠江水沙变化特征及人类活动影响因素分析[J]. 海洋学研究, 40(4): 52-64. |
杜庆顺, 黄慧, 吴晓晖, 等, 2022. 以广东省为例分析2021年冬2022年春珠江流域旱情及对策[J]. 中国防汛抗旱, 32(12): 89-92. |
李奕洁, 宋贵生, 胡素征, 等, 2015. 2014年夏季长江口有色溶解有机物(CDOM)的分布、光学特性及其来源探究[J]. 海洋与湖沼, 46(3): 670-678. |
李博, 蔡华阳, 杨昊, 等, 2023. 珠江河网横向汊道水位演变特性及原因探究[J]. 海洋与湖沼, 54(2): 313-330. |
吴增斌, 郭磊城, 吴雪枫, 等, 2022. 2020年特大洪水作用下长江口南槽水沙输移特征[J]. 海洋与湖沼, 53(2): 295-304. |
张先毅, 杨昊, 黄竞争, 等, 2020. 强人类活动驱动下珠江磨刀门河口径潮动力的季节性异变特征[J]. 海洋与湖沼, 51(5): 1043-1054. |
张连凯, 覃小群, 杨慧, 等, 2013. 珠江流域河流碳输出通量及变化特征[J]. 环境科学, 34(8): 3025-3034. |
张淑坤, 明玥, 高磊, 2022. 2020年夏季长江流域特大洪水期间长江口POC和DOC的分布特征[J]. 海洋环境科学, 41(5): 653-659. |
陈学秋, 杜勇, 付宇鹏, 2022. 2022年珠江流域暴雨洪水特点分析[J]. 中国水利, (22): 28-32. |
单森, 齐远志, 罗春乐, 等, 2020. 中国主要河流输送陆源碳的同位素特征及影响因素[J]. 地球科学进展, 35(9): 948-961. |
逄勇, 李毓湘, 2001. 珠江三角洲污染物对东四口门通量影响分析[J]. 河海大学学报, 29(4): 50-55. |
钱燕, 卢康明, 2022. 2021年珠江流域旱情分析与思考[J]. 中国防汛抗旱, 32(6): 27-30. |
钱燕, 卢康明, 陈学秋, 等, 2023. 珠江流域"2022.6"暴雨洪水复盘分析[J]. 中国防汛抗旱, 33(1): 22-26. |
曾丹娜, 牛丽霞, 陶伟, 等, 2020. 夏季珠江口水域营养盐分布特征及其富营养化评价[J]. 广东海洋大学学报, 40(3): 73-82. |
CALLAHAN J, DAI M H, CHEN R F, et al, 2004. Distribution of dissolved organic matter in the Pearl River Estuary, China[J]. Marine Chemistry, 89(1/2/3/4): 211-224. |
CHEN Z Q, LI Y, PAN J M, 2004a. Distributions of colored dissolved organic matter and dissolved organic carbon in the Pearl River Estuary, China[J]. Continental Shelf Research, 24(16): 1845-1856. DOI:10.1016/j.csr.2004.06.011 |
CHEN J F, LI Y, YIN K D, et al, 2004b. Amino acids in the Pearl River Estuary and adjacent waters: origins, transformation and degradation[J]. Continental Shelf Research, 24(16): 1877-1894. DOI:10.1016/j.csr.2004.06.013 |
COBLE P G, 1996. Characterization of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy[J]. Marine Chemistry, 51(4): 325-346. DOI:10.1016/0304-4203(95)00062-3 |
COBLE P G, 2013. Colored dissolved organic matter in seawater [M] // WATSON J, ZIELINSKI O. Subsea Optics and Imaging. Cambridge: Woodhead Publishing: 98-118.
|
DAI M, WANG L, GUO X, et al, 2008. Nitrification and inorganic nitrogen distribution in a large perturbed river/ estuarine system: the Pearl River Estuary, China[J]. Biogeosciences, 5(5): 1227-1244. DOI:10.5194/bg-5-1227-2008 |
GUO L D, MACDONALD R W, 2006. Source and transport of terrigenous organic matter in the upper Yukon River: Evidence from isotope (δ13C, Δ14C, and δ15N) composition of dissolved, colloidal, and particulate phases[J]. Global Biogeochemical Cycles, 20(2): GB2011. |
HE B Y, DAI M H, ZHAI W D, et al, 2010. Distribution, degradation and dynamics of dissolved organic carbon and its major compound classes in the Pearl River estuary, China[J]. Marine Chemistry, 119(1/2/3/4): 52-64. |
HELMS J R, STUBBINS A, RITCHIE J D, et al, 2008. Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter[J]. Limnology and Oceanography, 53(3): 955-969. DOI:10.4319/lo.2008.53.3.0955 |
HONG H S, WU J Y, SHANG S L, et al, 2005. Absorption and fluorescence of chromophoric dissolved organic matter in the Pearl River Estuary, South China[J]. Marine Chemistry, 97(1/2): 78-89. |
JOHNSTON S E, STRIEGL R G, BOGARD M J, et al, 2020. Hydrologic connectivity determines dissolved organic matter biogeochemistry in northern high‐latitude lakes[J]. Limnology and Oceanography, 65(8): 1764-1780. DOI:10.1002/lno.11417 |
KOTHAWALA D N, MURPHY K R, STEDMON C A, et al, 2013. Inner filter correction of dissolved organic matter fluorescence[J]. Limnology and Oceanography: Methods, 11(12): 616-630. DOI:10.4319/lom.2013.11.616 |
LI Y, SONG G S, MASSICOTTE P, et al, 2019. Distribution, seasonality, and fluxes of dissolved organic matter in the Pearl River (Zhujiang) estuary, China[J]. Biogeosciences, 16(13): 2751-2770. DOI:10.5194/bg-16-2751-2019 |
LIU Q Y, LIANG Y T, CAI W J, et al, 2020. Changing riverine organic C: N ratios along the Pearl River: Implications for estuarine and coastal carbon cycles[J]. Science of the Total Environment, 709: 136052. DOI:10.1016/j.scitotenv.2019.136052 |
LIU D, TIAN L Q, JIANG X T, et al, 2022. Human activities changed organic carbon transport in Chinese rivers during 2004-2018[J]. Water Research, 222: 118872. DOI:10.1016/j.watres.2022.118872 |
MANN P J, DAVYDOVA A, ZIMOV N, et al, 2012. Controls on the composition and lability of dissolved organic matter in Siberia's Kolyma River basin[J]. Journal of Geophysical Research: Biogeosciences, 117(G1): G01028. |
MING Y, GAO L, GUO L D, 2023. Dissolved and particulate organic carbon dynamics in the lower Changjiang River on timescales from seasonal to decades: Response to climate and human impacts[J]. Journal of Marine Systems, 239: 103855. DOI:10.1016/j.jmarsys.2022.103855 |
MURPHY K R, BUTLER K D, SPENCER R G M, et al, 2010. Measurement of dissolved organic matter fluorescence in aquatic environments: an interlaboratory comparison[J]. Environmental Science & Technology, 44(24): 9405-9412. |
MURPHY K R, STEDMON C A, GRAEBER D, et al, 2013. Fluorescence spectroscopy and multi-way techniques. PARAFAC[J]. Analytical Methods, 5(23): 6557-6566. DOI:10.1039/c3ay41160e |
PENG X Z, XIONG S S, OU W H, et al, 2017. Persistence, temporal and spatial profiles of ultraviolet absorbents and phenolic personal care products in riverine and estuarine sediment of the Pearl River catchment, China[J]. Journal of Hazardous Materials, 323: 139-146. DOI:10.1016/j.jhazmat.2016.05.020 |
SEITZINGER S P, MAYORGA E, BOUWMAN A F, et al, 2010. Global river nutrient export: A scenario analysis of past and future trends[J]. Global Biogeochemical Cycles, 24(4): GB0A08. |
SPENCER R G M, AIKEN G R, WICKLAND K P, et al, 2008. Seasonal and spatial variability in dissolved organic matter quantity and composition from the Yukon River basin, Alaska[J]. Global Biogeochemical Cycles, 22(4): GB4002. |
TAO Z, GAO Q Z, GUO W P, et al, 2011. Temporal and spatial variability of dissolved organic carbon concentration in the Xijiang River, South China[J]. Journal of Mountain Science, 8(5): 694-703. DOI:10.1007/s11629-011-2000-3 |
WANG Y, GAO L, 2022. Sources and dynamics of suspended particulate matter in a large-river dominated marine system: Contributions from terrestrial sediments, biological particles, and flocculation[J]. Journal of Marine Systems, 225: 103648. DOI:10.1016/j.jmarsys.2021.103648 |
WANG Y, GAO L, MING Y, et al, 2023. Recent declines in nutrient concentrations and fluxes in the Lower Changjiang River[J]. Estuaries and Coasts, 46(6): 1475-1493. DOI:10.1007/s12237-023-01216-8 |
WEI X, CAI S Q, NI P T, et al, 2020. Impacts of climate change and human activities on the water discharge and sediment load of the Pearl River, southern China[J]. Scientific Reports, 10(1): 16743. DOI:10.1038/s41598-020-73939-8 |
WEI X, WU C Y, 2014. Long-term process-based morphodynamic modeling of the Pearl River Delta[J]. Ocean Dynamics, 64(12): 1753-1765. DOI:10.1007/s10236-014-0785-7 |
WEISHAAR J L, AIKEN G R, BERGAMASCHI B A, et al, 2003. Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon[J]. Environmental Science & Technology, 37(20): 4702-4708. |
WORRALL F, BURT T P, 2007. Flux of dissolved organic carbon from U. K. rivers[J]. Global Biogeochemical Cycles, 21(1): GB1013. |
WU N, LIU S M, ZHANG G L, et al, 2021. Anthropogenic impacts on nutrient variability in the lower Yellow River[J]. Science of the Total Environment, 755: 142488. DOI:10.1016/j.scitotenv.2020.142488 |
XIA X M, LI Y, YANG H, et al, 2004. Observations on the size and settling velocity distributions of suspended sediment in the Pearl River Estuary, China[J]. Continental Shelf Research, 24(16): 1809-1826. DOI:10.1016/j.csr.2004.06.009 |
YAN L, XIE X J, PENG K, et al, 2021. Sources and compositional characterization of chromophoric dissolved organic matter in a Hainan tropical mangrove-estuary[J]. Journal of Hydrology, 600: 126572. DOI:10.1016/j.jhydrol.2021.126572 |
YANG S Y, TANG M, YIM W W S, et al, 2011. Burial of organic carbon in Holocene sediments of the Zhujiang (Pearl River) and Changjiang (Yangtze River) estuaries[J]. Marine Chemistry, 123(1/2/3/4): 1-10. |
ZHANG S R, LU X X, HIGGITT D L, et al, 2007. Water chemistry of the Zhujiang (Pearl River): natural processes and anthropogenic influences[J]. Journal of Geophysical Research: Earth Surface, 112(F1): F01011. |
ZHANG S R, LU X X, HIGGITT D L, et al, 2008. Recent changes of water discharge and sediment load in the Zhujiang (Pearl River) Basin, China[J]. Global and Planetary Change, 60(3/4): 365-380. |
ZHANG S Y, ZHANG H, 2023. Anthropogenic impact on long-term riverine CODMn, BOD, and nutrient flux variation in the Pearl River Delta[J]. Science of the Total Environment, 859: 160197. DOI:10.1016/j.scitotenv.2022.160197 |
ZHANG Y L, ZHOU Y Q, SHI K, et al, 2018. Optical properties and composition changes in chromophoric dissolved organic matter along trophic gradients: Implications for monitoring and assessing lake eutrophication[J]. Water Research, 131: 255-263. DOI:10.1016/j.watres.2017.12.051 |
ZHANG Y L, ZHOU L, ZHOU Y Q, et al, 2021. Chromophoric dissolved organic matter in inland waters: Present knowledge and future challenges[J]. Science of the Total Environment, 759: 143550. DOI:10.1016/j.scitotenv.2020.143550 |
ZHAO L B, GAO L, GUO L D, 2021. Seasonal variations in molecular size of chromophoric dissolved organic matter from the lower Changjiang (Yangtze) River[J]. Journal of Geophysical Research: Biogeosciences, 126(8): e2020JG006160. DOI:10.1029/2020JG006160 |
ZHOU Y Q, YAO X L, ZHOU L, et al, 2021. How hydrology and anthropogenic activity influence the molecular composition and export of dissolved organic matter: Observations along a large river continuum[J]. Limnology and Oceanography, 66(5): 1730-1742. DOI:10.1002/lno.11716 |