 2024, Vol. 55
2024, Vol. 55中国海洋湖沼学会主办。
文章信息
- 李子东, 汤建华, 吴磊, 闫欣, 葛慧, 施金金, 王燕平, 朱海晨, 王储庆. 2024.
- LI Zi-Dong, TANG Jian-Hua, WU Lei, YAN Xin, GE Hui, SHI Jin-Jin, WANG Yan-Ping, ZHU Hai-Chen, WANG Chu-Qing. 2024.
- 长江禁捕前江苏海域刀鲚(Coilia nasus)时空分布及环境因子的初步研究
- SPATIO-TEMPORAL DISTRIBUTION AND INFLUENCING FACTORS OF COILIA NASUS IN SEA AREAS OFF JIANGSU BEFORE FISHING BAN IN CHANGJIANG RIVER
- 海洋与湖沼, 55(2): 441-450
- Oceanologia et Limnologia Sinica, 55(2): 441-450.
- http://dx.doi.org/10.11693/hyhz20231100239
文章历史
-
收稿日期:2023-11-13
收修改稿日期:2023-12-29




刀鲚(Coilia nasus), 俗名刀鱼、凤尾鱼, 学名长颌鲚, 隶属于鲱形目(Clupeiformes)、鳀科(Engraulidae)、鲚属(Coilia)。刀鲚在我国分淡水定居和江海洄游两种生态类型, 其中江海洄游类群分布广泛, 从南到北在钱塘江、长江、淮河、海河、辽河及其他的通海河流中均有发现, 且以长江流域产量为最高(袁传宓等, 1984; 贺刚等, 2017)。每年春季(2月下旬至3月初), 性成熟的刀鲚在长江口集成大群, 由海洋入长江作生殖洄游, 产卵后再返回近海育肥。作为“长江三鲜”之一, 刀鲚因味道鲜美在市场拥有较高的认可度, 又因能形成稳定渔汛, 成为沿江各省渔业主要利用经济鱼类。20世纪50年代末至70年代初, 长江刀鲚产量一直处于上升状态(袁传宓, 1988), 20世纪70~80年代, 上海市、江苏省及安徽省刀鲚年均捕捞量为2 904 t, 其中1973年最高, 约为4 142 t (代培等, 2020)。20世纪90年代后, 由于受水环境污染加重、持续高强度捕捞、幼鱼资源遭到破坏、水利工程阻隔洄游等因素影响, 刀鲚产量出现大幅下滑。2001~2005年刀鲚平均捕捞量急剧下降至673 t, 相比于20世纪70年代, 降幅为76.83% (施炜纲等, 2009); 2010~2013年平均捕捞量进一步下降, 为116.6 t, 其相比于20世纪90年代, 降幅为86.40%, 相比于20世纪70年代, 降幅达95.98%, 渔汛濒临消失的窘境(徐跑等, 2016)。姜涛等(2022)基于2014~2020年在鄱阳湖及周边水域的连续调查监测, 发现长江禁渔前(2014~2018年)溯河洄游型刀鲚单位捕捞努力量渔获量(CPUE)仅为(0.81±1.73)×10–5 ind./(h·m), 即(1.13±2.42) ind./d; 而在禁渔实施两年后(2019~2020年), 该水域刀鲚CPUE已提升至(66.14±69.74)×10–5 ind./(h·m), 即(92.60± 97.64) ind./d, 前后对比增长了约82倍(姜涛等, 2022)。
以往, 学者对刀鲚的研究多聚焦在长江口、长江中下游江段及支流湖泊, 研究方向包括生物学与遗传特征(张媛等, 2006; 张堂林等, 2008; 杨彦平等, 2021)、年龄结构与生长特征(郭弘艺等, 2006; 黎雨轩等, 2010; 郑飞等, 2012; 罗宇婷等, 2021)、地理种群判别(程起群等, 2004; 王丹婷等, 2012; 张赛赛等, 2020; 李宇等, 2022a; Zhang et al, 2023)、资源量与群体结构(万全等, 2009; 刘凯等, 2012; 郭弘艺等, 2016)、基因组学与信号通路研究(Zhang et al, 2016; Liu et al, 2019; Wang et al, 2020)、生境履历重建(陈婷婷等, 2016; 李孟孟等, 2017; 李宇等, 2022b)等。但刀鲚在淡水水域产卵后有返回近海育肥的习性, 目前学界对刀鲚在海洋中的生物学特征及地理分布缺乏系统性研究已成为探究刀鲚生命规律的一大短板。江苏海域紧邻长江口, 自然生态条件优越, 是刀鲚亲鱼育肥与幼鱼成长的理想生境, 也是长江流域刀鲚传统栖息海域。因此, 分析江苏海域刀鲚的规格, 探究其地理分布情况, 有利于更加全面的认识长江刀鲚生命规律, 从而为该流域刀鲚种群的科学评估与合理利用提供数据支持。2020年, 农业农村部发布《农业农村部关于设立长江口禁捕管理区的通告》, 设立长江口禁捕管理区, 规定自2021年1月1日起长江口禁捕管理区以内水域, 实行长江流域禁捕管理制度。在此背景下, 江苏海域涵盖了长江刀鲚种群近岸活动海域与长江口产卵海域, 在该海域进行渔业资源调查, 既能完善长江刀鲚生活史, 也能为后期长江禁渔效果的科学评估提供第一手佐证材料。
1 材料与方法 1.1 调查设计 1.1.1 调查海域范围、时间本研究数据来源于江苏省水生生物资源重大专项暨首次水生野生动物资源普查-近海海洋水生野生动物资源普查, 调查区域为江苏海域(31°45′~35°00′N, 119°30′~125°00′E), 包括江苏近岸海域(江苏境内“禁渔区线”到海岸线之间水域)与江苏境内“禁渔区线”外侧水域, 南黄海辐射沙脊群也位于本次调查海域内。调查时间分为2017年春季(5月)、夏季(8月)、秋季(11月)和冬季(2018年2月)共4个季度。
1.1.2 调查站位设置本研究共设置68个定点调查站位, 其中“禁渔区线”内侧海域29个站位(JS44站位划归“禁渔区线”内侧海域), “禁渔区线”外侧海域39个站位。“禁渔区线”内侧基本按照每30′一个站点设置, 因复杂的水文状况与辐射沙脊群独有的特殊生境, 部分站点设置略有调整。“禁渔区线”外侧则按照每隔经纬度30′设置一个站点均匀布设(图 1)。
 |
| 图 1 江苏海域调查站位图 Fig. 1 Sketch map of the sampling stations in sea areas off Jiangsu |
本次调查在“禁渔区线”内、外两侧同步进行, 四个季度“禁渔区线”内调查船均为苏通渔01026号, 吨位140 t, 其主机功率220 kW, 调查渔具为单船有翼单囊拖网, 网口拉紧周长90 m, 扫海宽度17.5 m, 网囊网目25 mm; “禁渔区线”外调查船均为浙嵊渔10201号和浙嵊渔10243号, 吨位150 t, 其主机功率280 kW, 调查渔具为双船有翼单囊拖网, 网口拉紧周长400 m, 扫海宽度35.0 m, 网囊网目25 mm, 调查渔船拖速均在2.0~3.0 kn, 每个站位原则上拖曳1 h。
1.2 研究方法 1.2.1 采样和分析方法海上调查以及生物学测定根据海洋调查规范(GB/T-12763.6-2007) (中华人民共和国国家质量监督检验检疫总局等, 2008)进行操作, 渔获采样放网位置要综合拖速、拖向等多种因素确定, 在距标准站点2~4 n mile时放网, 确保经1 h拖网后正好到标准站位位置或附近, 拖网过程中要确保拖网方位朝向正确, 起网前须准确测定船位(国家海洋局908专项办公室, 2006), 捕捞作业时现场记录起放网时间、起放网经纬度、渔获量等各项渔捞要素。
现场采集的样本冰鲜后保存在实验室冰柜, 解冻后进行生物学测定, 先鉴定挑选出刀鲚样品(样本抽样见表 1), 再以海洋调查规范(GB/T-12763.6-2007)(中华人民共和国国家质量监督检验检疫总局等, 2008)为标准测定全长(BL)并精确至1 mm, 体重(BW)精确至0.1 g。本文选取的影响刀鲚分布环境因素包括水温、盐度、水深、溶解氧、透明度, 在本调查中透明度采用塞氏盘SD20获取, 其他环境因子则用Ocean seven-304型CTD获取。
| 季节 | 捕获样本/尾 | 抽样样本/尾 | 抽样占比/% |
| 春季 | 70 | 51 | 72.9 |
| 夏季 | 468 | 468 | 100.0 |
| 秋季 | 2 713 | 658 | 24.3 |
| 冬季 | 2 321 | 1 821 | 78.5 |
| 总计 | 5 572 | 2 998 | 53.8 |
本研究先将样品计数(尾)和称重(kg), 再将尾数和重量转换为尾数资源密度(ind./km2)和重量资源密度(kg/km2), 对单船有翼单囊拖网和双船有翼单囊拖网的调查获得的数据进行了标准化处理, 以尾数资源密度和重量资源密度为指标分析刀鲚在江苏海域四季时空分布规律。其计算公式如下(郑元甲等, 2003):
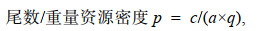 (1)
(1)式中, p为资源密度(ind./km2, kg/km2); c为平均每小时拖网渔获量[ind./(net∙h), kg/(net∙h)]; a为网具每小时扫海面积[km2/(net∙h)]; q为网具捕获率(即捕捞系数, 刀鲚属暖温性中上层鱼类, q取值0.3) (水柏年, 2017; 王尽文等, 2018)。
根据全长、体重直方图数据将全长、体重组距值分别定为50 mm、30 g, 绘制全长、体重频率分布图。数据分析和处理利用Excel 2019及SPSS 26完成, 尾数资源密度及重量资源密度分布图利用Arcmap 10.2软件绘制。采用SPSS 10.2软件拟合刀鲚全长与体质量之间关系: W=aLb, 其中, W为体质量(g), L为全长(mm), a、b为参数, a为生长的条件因子, b为幂指数函数(詹秉义, 2005)。同时, 配对t检验确定参数b与3的显著性差异(马凤娇等, 2022)。
相关分析是统计分析方法中最重要的内容之一, 也是多元统计分析方法的基础。当分析多个事物间的关系, 且这种关系又呈现为变量间的数量关系时, 可以采用双变量相关分析方法, 其输出的统计量包括均值、标准差、Spearman相关系数、协方差阵等(林则宏等, 2017)。本研究结合刀鲚生物学特性与生态类型(暖温性、中上层鱼类)筛选出影响刀鲚生境的五个主要环境因子: 深度(m)、温度(℃)、盐度、溶解氧(×10–6)、透明度(m), 利用双变量相关分析中的Spearman检验研究调查站点刀鲚生物量与其主要环境因子之间的关系, 双变量相关分析(Spearman检验)使用IBM SPSS Statistics 26软件进行。
2 结果 2.1 全长频率分布刀鲚全长的全年变幅为46~362 mm, 平均值为(135.37±37.27) mm, 春季刀鲚全长平均值为(143.79± 28.26) mm, 夏季刀鲚全长平均值为(138.79±8.16) mm, 秋季刀鲚全长平均值为(157.75±14.65) mm, 冬季刀鲚全长平均值为(128.45±9.60) mm。各季节的全长频率占比分布如图 2所示, 江苏海域四个季节刀鲚样本的全长优势组均为101~150 mm, 呈夏季占比(71.20%) > 春季(68.97%) > 冬季(63.45%) > 秋季(54.20%)的趋势; 全长范围为51~100 mm的小规格刀鲚与151~200 mm组的较大规格刀鲚也占据较大比例, 分别呈现出冬季占比(16.63%) > 夏季(5.76%) > 春季(3.45%) > 秋季(0.87%)、秋季(31.59%) > 春季(24.14%) > 冬季(16.79%) > 夏季(14.55%)的趋势; 全长范围为201~250 mm的大规格刀鲚全长占比则为秋季占比(8.70%) > 春季(8.38%) > 冬季(1.69%) > 夏季(0.00%); 全长在251~300 mm范围的刀鲚占比较小, 为秋季(4.06%) > 春季(3.45%) > 冬季(1.29%) > 夏季(0.00%); 全长在301~ 350 mm范围的刀鲚样本占比很小, 样本仅在秋季(0.29%)与冬季(0.08%) 在研究海域出现; 全长在351~400 mm范围的刀鲚样本占比极小, 仅在秋季(0.29%)收集到极少数样本。
 |
| 图 2 江苏海域四季刀鲚全长分布 Fig. 2 Body length distribution of C. nasus in various season in Jiangsu sea area |
刀鲚体重的全年变幅为0.3~153.2 g, 平均值为(9.77±10.59) g, 春季刀鲚体重平均值为(11.12±14.53) g, 夏季刀鲚体重平均值为(9.13±8.16) g, 秋季刀鲚体重平均值为(14.03±14.65) g, 冬季刀鲚体重平均值为(9.01±9.60) g。各季节的体重频率占比分布如图 3所示, 江苏海域四个季节刀鲚样本体重绝对优势组均为0~30 g, 呈春季占比(96.55%) > 夏季(94.76%) > 秋季(88.99%) > 秋季(67.20%)的趋势; 体重范围在31~60 g区间的刀鲚样本占比次之, 呈秋季占比(9.28%) > 夏季(5.24%) > 冬季(0.94%) > 春季(0.00%)趋势; 体重范围在61~90 g区间的刀鲚样本占比更少, 呈春季(3.45%) > 秋季(1.16%) > 冬季(0.44%) > 夏季(0.00%); 体重范围在91~120 g的刀鲚样本在秋季(0.29%)与冬季(0.06%)有极少占比; 体重范围在151~180 g的刀鲚样本仅在秋季(0.29%)有极少占比。
 |
| 图 3 江苏海域四季刀鲚体重分布 Fig. 3 Body weight distribution of C. nasus in four seasons in sea areas off Jiangsu |
依据2017年5月~2018年2月江苏海域刀鲚全长L (单位: mm)与体质量W (单位: g)数据, 在不考虑雌雄差异的情况下, 拟合江苏海域刀鲚全长-体质量的幂函数关系式(图 4), 其关系式为:
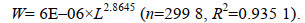 (2)
(2)
 |
| 图 4 江苏海域刀鲚全长与体质量关系 Fig. 4 The relationship between body length and body weight of C. nasus in sea areas off Jiangsu |
配对t检验结果表明, 参数b与3存在极显著差异(P < 0.01), 且b < 3, 即本次调查实施期间(2017年5月~2018年2月)江苏海域刀鲚呈现负异速生长趋势。
2.4 资源密度的季节变化江苏海域刀鲚尾数资源密度冬季(2月)最高, 为3 211.3 ind./km2; 秋季(11月)次之, 为1 714.6 ind./km2; 其余依次为夏季(8月)和春季(5月), 分别为89.3和15.6 ind./km2。
重量资源密度仍以冬季最高20.1 kg/km2; 秋季次之, 16.8 kg/km2; 其他依次为夏季和春季, 分别为0.6和0.2 kg/km2 (表 2)。
| 类别 | 生物密度 | 春季 | 夏季 | 秋季 | 冬季 |
| 尾数资源密度/(ind./km2) | 最低 | 0.0 | 0.0 | 0.0 | 0.0 |
| 最高 | 408.2 | 3 200.0 | 71 059.3 | 42 315.0 | |
| 平均 | 15.6 | 89.3 | 1 747.6 | 3 211.3 | |
| 重量资源密度/(kg/km2) | 最低 | 0.0 | 0.0 | 0.0 | 0.0 |
| 最高 | 3.8 | 17.9 | 550.3 | 266.6 | |
| 平均 | 0.2 | 0.6 | 16.8 | 20.1 |
由图 5可知, 从区域变化来看, 春季, 江苏海域刀鲚集中分布于“禁渔区线”内侧的长江口以北海域, 外侧海域未收集到刀鲚样本, 尾数资源密度中心分布区位于32°45′~33°15′N, 121°15′~121°45′E, 另有JS24、JS32、JS62站位周边海域的尾数资源密度也较高; 夏季, 刀鲚分布仍在“禁渔区线”内侧, 尾数资源密度中心分布区相比春季向北延伸, 资源密度中心分布区东西分布界限为“禁渔区线”, 南北分布界限为32°30′~ 34°30′N; 秋季, 刀鲚尾数资源密度中心分布区开始移至“禁渔区线”外侧的33°0′~34°30′N, 121°30′~ 122°0′E海域, “禁渔区线”内侧南部的JS60、JS61、JS62、JS63、JS68站位的尾数资源密度也相对较高; 冬季, 刀鲚尾数资源密度中心区相比秋季南移趋势显著, 且绝大部分站位均位于“禁渔区线”内侧长江口北部海域(31°45′~32°30′N, 121°45′~122°30′E), 外侧仅有JS25、JS35、JS45、JS54、JS64、JS65等6个站位的尾数资源密度较高。
 |
| 图 5 江苏海域刀鲚四季尾数资源密度分布 Fig. 5 The horizontal distribution of density index of stock of C. nasus in four seasons in sea areas off Jiangsu 注: a. 春季; b. 夏季; c. 秋季; d. 冬季 |
由图 6可知, 春季, 江苏海域刀鲚的重量资源密度中心分布区位于JS42、JS50、JS62、JS24站位周边海域, 重量资源量为全年最小; 夏季, 重量资源量相比春季变大, 且刀鲚的重量资源密度中心分布区位于32°30′~34°30′N、“禁渔区线”内侧的江苏近岸海域, 其分布相比春季有明显的偏北趋势; 秋季, 刀鲚重量资源密度中心分布区转移至“禁渔区线”外侧33°0′~34°30′N, 121°30′~122°0′E的小块海域, “禁渔区线”内侧南部的JS50、JS60、JS63站位重量资源密度也较高, 秋季重量资源密度值相比夏季进一步增大; 冬季, 刀鲚重量资源密度达到全年最大值, 34°N线以南、“禁渔区线”内外两侧均有较高重量资源密度站位, 重量资源密度中心分布区相比秋季同样呈现南移趋势, 多数站位均位于长江口北部海域(31°45′~32°30′N, 121°45′~122°30′E)。
 |
| 图 6 江苏海域刀鲚四季重量资源密度分布 Fig. 6 The horizontal distribution of density index of weight of C. nasus in four seasons in sea areas off Jiangsu 注: a. 春季; b. 夏季; c. 秋季; d. 冬季 |
为探明不同季节影响江苏海域刀鲚生物量的环境因子, 将四季不同站位的刀鲚生物量分别与该海域采集的表层、底层环境因子做双变量相关性分析(Spearman检验), 见表 3。结果表明: 江苏海域春季刀鲚生物量与表层温度(R=0.253, P=0.037)、表层溶氧(R=0.265, P=0.029)、透明度(R=–0.307, P=0.011)、水深(R=–0.272, P=0.025)、底层温度(R=–0.291, P=0.016)、底层盐度(R=–0.306, P=0.011)存在显著相关关系, 与表层盐度之间存在极显著相关性(R=–0.422, P=0.000); 夏季刀鲚生物量与透明度存在显著相关性(R=–0.235, P=0.037), 与其余8个环境因子无显著相关性; 秋季刀鲚生物量与全部9个环境因子之间均无显著相关性(P > 0.05); 冬季刀鲚生物量与水深之间存在显著相关性(R=–0.252, P=0.038), 与其余8个环境因子无显著相关性。
| 环境因子 | 春季 | 夏季 | 秋季 | 冬季 |
| 表层温度 | R=0.253 | R=–0.037 | R=0.018 | R=–0.199 |
| P=0.037 | P=0.766 | P=0.887 | P=0.103 | |
| 表层盐度 | R=–0.422 | R=–0.024 | R=0.062 | R=–0.107 |
| P=0.000 | P=0.844 | P=0.617 | P=0.386 | |
| 表层溶氧 | R=0.265 | R=0.057 | R=0.110 | R=0.149 |
| P=0.029 | P=0.644 | P=0.373 | P=0.225 | |
| 透明度 | R=–0.307 | R=–0.254 | R=–0.009 | R=–0.161 |
| P=0.011 | P=0.037 | P=0.941 | P=0.190 | |
| 水深 | R=–0.272 | R=–0.235 | R=–0.149 | R=–0.252 |
| P=0.025 | P=0.054 | P=0.226 | P=0.038 | |
| 底层温度 | R=–0.291 | R=0.144 | R=0.134 | R=–0.161 |
| P=0.016 | P=0.242 | P=0.275 | P=0.189 | |
| 底层盐度 | R=–0.306 | R=–0.060 | R=–0.066 | R=–0.040 |
| P=0.011 | P=0.628 | P=0.594 | P=0.747 | |
| 底层溶氧 | — | R=–0.073 | R=0.060 | R=0.136 |
| — | P=0.557 | P=0.625 | P=0.267 | |
| 注: 春季底层溶氧环境因子数据缺失, 用“—”表示 | ||||
结合前人在长江中下游水域(包括各通江河道和湖泊)即长江口对刀鲚产卵洄游习性研究结果与江苏海域刀鲚全长、体重分布, 资源密度大小情况, 解析该海域刀鲚四季时空分布情况。资料记载, 性成熟的刀鲚会在每年2~9月分批从海洋进入长江, 上溯至产卵场进行产卵繁殖(刘引兰等, 2008)。春季(5月), 江苏海域刀鲚尾数资源密度与重量资源密度均为全年最低值, 这可能是由于在本研究取样时刀鲚已进入产卵旺季, 大量的刀鲚成鱼进入长江溯游产卵, 导致近岸海域刀鲚数量急剧减少。历史研究表明, 20世纪80、90年代长江刀鲚的产卵场最远可到达距离长江口800 km的鄱阳湖以及1 400 km的洞庭湖一带(袁传宓等, 1980; 朱栋良, 1992)。21世纪以来, 由于长江流域内刀鲚栖息环境持续恶化, 加之水利工程的建设和过度捕捞, 使得刀鲚很难溯流洄游到洞庭湖与鄱阳湖等水域产卵(廖伏初等, 2002)。姜涛等(2013)、轩中亚等(2020)、Sokta等(2020)基于耳石微化学对鄱阳湖、洞庭湖内刀鲚的生境履历进行研究, 发现这两个历史上长江刀鲚的传统产卵场既存在溯河洄游型刀鲚, 也存在淡水定居型刀鲚, 目前长江流域刀鲚产卵场具体分布仍存在疑团, 需要后来研究者探索解秘。夏季(8月), 刀鲚尾数资源密度与重量资源密度相比春季均有增大, 这是因为较早出发溯游产卵的刀鲚成鱼已在淡水产卵完毕, 与当年生幼鱼从长江返回江苏近海休养索饵, 其尾数与重量资源密度中心分布区(33°0′~34°30′N, 120°30′~121°30′E)相比春季更加偏北, 均分布于江苏近岸海域; 秋季(11月), 随着返海刀鲚数量增加, 江苏海域刀鲚的尾数资源密度与重量资源密度相比夏季大幅增长, 加之此季节水温下降, 秋季尾数与重量资源密度中心分布区(33°0′~ 34°30′N, 121°30′~122°0′E)开始向“禁渔区线”外侧的深海区移动, 刀鲚逐渐进入越冬期; 刀鲚在性腺最终成熟前沿长江迁徙至邻近湖泊产卵, 产卵期通常为2~4月(朱春月等, 2023), 故在冬季(2月)时, 江苏海域刀鲚逐渐在长江口集成大群, 准备新一轮的产卵洄游, 其尾数与重量资源密度中心分布(31°45′~32°30′N, 121°45′~ 122°30′E)相比秋季明显南移, 集中分布于长江口附近海域。
3.2 影响江苏海域刀鲚时空分布的环境因子以往研究发现, 2月初的江苏省江段即可见到上溯准备产卵的刀鲚鱼群, 3、4月份长江口附近的刀鲚鱼群达到高潮阶段, 称为刀鲚汛期(袁传宓, 1987), 本研究中春季调查实施月份(5月)正值刀鲚产卵旺期。而刀鲚进入性成熟阶段后, 性腺分泌的激素会引起鱼体内一系列变化, 如渗透压的改变, 开始出现产卵环境选择行为等, 为寻找拥有适宜盐度、水温、流速等的产卵环境, 刀鲚开始生殖洄游(袁传宓, 1987)。淡水水域渗透压低于海水, 既适宜鱼卵的孵化, 同时湖湾、水湾的水面平静, 又能提供丰富的饵料, 成为刀鲚产卵的最佳选择。本研究发现, 表层温度(R=0.253, P=0.037)、表层溶氧(R=0.265, P=0.029)与春季江苏海域刀鲚分布存在显著正相关关系, 而透明度(R=–0.307, P=0.011)、水深(R=–0.272, P=0.025)、底层温度(R=–0.291, P=0.016)、底层盐度(R=–0.306, P=0.011)则与刀鲚分布存在显著负相关关系。这可能与长江刀鲚在海洋不集成大群, 多分散在沿海及附近海域的底层, 属近海底层鱼类, 到进入长江口附近才会逐渐集成大群, 栖息水层也由底层上升到中、上层的生态习性有关(刘引兰等, 2008; 田思泉等, 2014)。同时, 处于生殖洄游期间的刀鲚对能量的需求较高, 而水温的高低将直接影响其生理和代谢活动, 水温升高能够加速刀鲚的性腺发育(马凤娇等, 2022), 故刀鲚洄游受温度的影响较大。夏季(8月), 部分较早溯河洄游的刀鲚已产卵结束, 开始降河洄游至江苏近岸海域索饵育肥, 出现“回头刀鱼”现象。此时刀鲚刚结束产卵不久, 返回近海后急需进食补充能量, 丰富的营养物质成为影响其分布的主要因素。从生存策略来看, 一方面, 刀鲚产卵时多喜栖息在水流混浊的水域中, 清水中较少(刘引兰等, 2008), 另一方面, 刀鲚在沿长江产卵洄游期间一般不进食, 主要依靠体内积累的营养物质完成整个溯河洄游生殖过程, 直到它们迁徙到产卵地点才开始进食(姜涛等, 2018)。刀鲚的这些生存策略既从侧面印证了饵料对产卵后刀鲚分布的影响, 也与江苏海域夏季刀鲚分布与营养物质丰富程度存在显著相关关系的结论相印证; 从水文特征来看, 调查实施月份(8月)为北半球夏季, 正值江苏境内60余条入海河流的汛期, 此时长江、淮河、灌河、通榆河等大河入海水量显著增加, 带来更多的营养物质与泥沙, 浮游生物在此聚集。加之该海域内苏北沿岸锋面的影响作用(刘东艳等, 2022), 江苏近岸水质更加肥沃, 透明度变低, 形成高生产力海域, 吸引着急需补充能量的返海刀鲚前来索饵; 秋季(11月), 江苏海域水温下降, 刀鲚作为暖温性鱼类, 对水温变化敏感, 开始向外海洄游以寻找生境适宜的越冬场越冬。本研究环境数据来自2017~2018一年的资源调查, 相关性分析结果具有一定的偶然性, 未发现秋季刀鲚分布与水温之间存在相关性, 但结合刀鲚生态习性(暖温性底层鱼类)及秋季资源密度分布规律(高资源密度区集中在“禁渔区线”外侧海域), 作者推测水温是影响秋季刀鲚分布的重要因子, 这一观点需要后续研究者在该海域进行连续观测加以验证; 冬季(2月), 刀鲚性腺再次发育, 逐渐在长江口附近聚集成大群, 新一轮的生殖洄游开始, 此时刀鲚栖息水层从底层上升到中、上层, 反映在环境因子上, 水深与刀鲚分布之间呈显著负相关关系(R=–0.252, P=0.038), 即水深越深, 刀鲚生物量越少。
3.3 禁捕管理对江苏海域刀鲚的影响为保护长江流域刀鲚资源, 2018年12月28日农业农村部发布《农业农村部关于调整长江流域专项捕捞管理制度的通告》(中华人民共和国农业农村部, 2018), 规定自2019年2月1日起, 停止发放刀鲚专项捕捞许可证, 禁止刀鲚天然资源的生产性捕捞。2020年11月20日, 农业农村部又发布了《农业农村部关于设立长江口禁捕管理区的通告》(中华人民共和国农业农村部, 2020), 决定扩延长江口禁捕范围, 设立长江口禁捕管理区(122°15′E、31°41′36″N、30°54′N形成的框型区线, 向西以水陆交界线为界), 自2021年1月1日0时长江口禁捕管理区以内水域, 实行长江流域禁捕管理制度, 这一规定为长江刀鲚资源的保护与后续恢复奠定了制度基石。渔获规格通常是生物学最小型的直接表现, 能在一定程度上反映某一种群的变化趋势。万全等(2009)于2006年3~7月在安徽无为长江段对刀鲚生殖洄游群体进行了资源调查, 发现最高网次捕获量仅0.85 kg, 抽取100尾刀鲚测定, 发现体长为217~421 mm、体重为20~235 g; 郑飞等(2012)分析了2009年4~5月在长江九段沙、靖江和芜湖3个江段采集的溯河洄游型刀鲚样本, 发现3个种群的体长变幅为158~328 mm, 平均体长为(233±35) mm; 体重变幅为11.8~143.8 g, 平均体重为(48.19±24.89) g。本研究发现2017年5月~2018年2月江苏海域刀鲚全长变幅为46~362 mm, 平均值为(135.37±37.27) mm, 优势组全长为109~172 mm; 体重变幅为0.3~153.2 g, 平均值为(9.77± 10.59) g。对比以往调查资料, 可知2006~2018年间长江流域的洄游型刀鲚的体长和体重均显著减小, 规格变化明显。再结合马凤娇等(2022)在2019~2021年长江口持续调查的研究结果, 发现自2019年专项捕捞退出后, 长江刀鲚的体长、体重开始逐渐增长, 刀鲚体长和体质量显著大于本研究中2017~2018年“水野普查”时调查数据, 这表明随着2019年刀鲚生产性捕捞的退出, 2020年长江流域重点水域全面禁捕的实施和长江口禁捕管理区的成立, 长江刀鲚资源呈恢复趋势。可见, 长江十年禁渔的全面实施, 极大地缓解了渔业资源过度捕捞压力, 使得长江流域包括刀鲚在内的众多水生物种得到了休养恢复。但考虑到长江沿岸的人类生产生活的干扰、水利水电设施的影响等仍将长期存在, 建议对刀鲚等洄游性旗舰物种开展长期性、常态化监测, 从而能更加科学、系统地评估长江十年禁渔效果与流域内的渔业资源恢复状况。
4 结论本研究突破了以往刀鲚资源调查多在长江内水与河口的地理限制, 分析发现长江刀鲚入海后的大部分生命周期均在江苏近岸海域内栖息洄游, 即江苏近岸海域是其主要栖息索饵水域; 春、夏、秋、冬四季影响江苏海域刀鲚分布的环境因子各不相同, 而尤以春季影响因子最多; 对比长江禁捕后收集到的刀鲚样本, 发现“长江禁捕”实施后刀鲚种群渔获规格逐渐增大, 全长、体重逐渐增长, 刀鲚种群呈现出资源恢复趋势。
万全, 赖年悦, 李飞, 等, 2009. 安徽无为长江段刀鲚生殖洄游群体年龄结构的变化分析[J]. 水生态学杂志, 30(4): 60-65. |
马凤娇, 杨彦平, 方弟安, 等, 2022. 长江禁捕后长江口刀鲚资源特征[J]. 水生生物学报, 46(10): 1580-1590. |
王丹婷, 杨健, 姜涛, 等, 2012. 不同水域刀鲚形态的分析比较[J]. 水产学报, 36(1): 78-90. |
王尽文, 陶卉卉, 张乃星, 等, 2018. 2016年春季日照港岚山港区近岸海域渔业资源浅析[J]. 广西科学院学报, 34(2): 125-129, 136. |
中华人民共和国农业农村部, 2018. 农业农村部关于调整长江流域专项捕捞管理制度的通告[EB/OL]. (2018-12-28) [2018-12-28]. http://www.cjyzbgs.moa.gov.cn/zcjd/201905/t20190521_6313317.htm.
|
中华人民共和国农业农村部, 2020. 农业农村部关于设立长江口禁捕管理区的通告[EB/OL]. (2020-11-20) [2020-11-20]. http://www.cjyzbgs.moa.gov.cn/tzgg/202011/t20201120_6356569.htm.
|
中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会, 2008. 海洋调查规范第1部分: 总则: GB/T 12763.1-2007[S]. 北京: 中国标准出版社.
|
水柏年, 2017. 渔业资源调查与评价[M]. 北京: 海洋出版社, 40.
|
田思泉, 田芝清, 高春霞, 等, 2014. 长江口刀鲚汛期特征及其资源状况的年际变化分析[J]. 上海海洋大学学报, 23(2): 245-250. |
代培, 严燕, 朱孝彦, 等, 2020. 长江刀鲚国家级水产种质资源保护区(安庆段)刀鲚资源现状[J]. 中国水产科学, 27(11): 1267-1276. |
朱春月, 杨培民, 2023. 刀鲚洄游生物学及应激胁迫研究进展[J]. 中国农学通报, 39(2): 130-134. |
朱栋良, 1992. 长江刀鱼的天然繁殖与胚胎发育观察[J]. 水产科技情报, 19(2): 49-51. |
刘引兰, 吴志强, 胡茂林, 2008. 我国刀鲚研究进展[J]. 水产科学, 27(4): 205-209. |
刘东艳, 吕婷, 林磊, 等, 2022. 我国近海陆架锋面与生态效应研究回顾[J]. 海洋科学进展, 40(4): 725-741. |
刘凯, 段金荣, 徐东坡, 等, 2012. 长江口刀鲚渔汛特征及捕捞量现状[J]. 生态学杂志, 31(12): 3138-3143. |
李宇, 冯广朋, 张涛, 等, 2022a. 6个不同水域刀鲚的形态特征与聚类分析[J]. 水生态学杂志, 43(6): 116-123. |
李宇, 陈建华, 冯广朋, 等, 2022b. 基于耳石微化学的刀鲚群体生境履历研究进展[J]. 渔业信息与战略, 37(3): 196-203. |
李孟孟, 姜涛, KHUMBANYIWA D D, 等, 2017. 基于耳石微化学的长江安徽和县江段刀鲚生境履历重建[J]. 水生生物学报, 41(5): 1054-1061. |
杨彦平, 许萌原, 马凤娇, 等, 2021. 基于线粒体Cyt b基因的长江刀鲚群体遗传结构分析[J]. 江西农业学报, 33(8): 11-16, 23. |
轩中亚, 姜涛, 刘洪波, 等, 2020. 洞庭湖中是否存在溯河洄游型刀鲚[J]. 水生生物学报, 44(4): 838-843. |
张堂林, 李钟杰, 曹文宣, 2008. 鱼类生态形态学研究进展[J]. 水产学报, 32(1): 152-160. |
张媛, 胡则辉, 周志刚, 等, 2006. 利用RAPD-PCR与ISSR-PCR标记技术分析长江口刀鲚的群体遗传结构[J]. 上海水产大学学报, 15(4): 390-397. |
张赛赛, 罗珺, 陈博锦, 等, 2020. 不同群体刀鲚形态差异研究[J]. 淡水渔业, 50(3): 26-33. |
陈婷婷, 姜涛, 李孟孟, 等, 2016. 长江南京江段长颌鲚生境履历的反演[J]. 水产学报, 40(6): 882-892. |
林则宏, 段梅素, 程红英, 2017. 统计学[M]. 上海: 上海交通大学出版社, 106-109.
|
国家海洋局908专项办公室, 2006. 海洋生物生态调查技术规程[M]. 北京: 海洋出版社, 250.
|
罗宇婷, 代培, 刘思磊, 等, 2021. 2018年长江安庆段洄游汛期刀鲚年龄结构和生长特征[J]. 广东海洋大学学报, 41(3): 36-43. |
郑飞, 郭弘艺, 唐文乔, 等, 2012. 溯河洄游的长江刀鲚种群的年龄结构及其生长特征[J]. 动物学杂志, 47(5): 24-31. |
郑元甲, 陈雪忠, 程家骅, 等, 2003. 东海大陆架生物资源与环境[M]. 上海: 上海科学技术出版社, 53-57.
|
施炜纲, 张敏莹, 刘凯, 等, 2009. 水工工程对长江下游渔业的胁迫与补偿[J]. 湖泊科学, 21(1): 10-20. |
姜涛, 刘洪波, 李孟孟, 等, 2018. 溯河洄游长江刀鲚(Coilia nasus)摄食虾类的调查[J]. 湖泊科学, 30(2): 458-463. |
姜涛, 杨健, 轩中亚, 等, 2022. 长江禁渔对鄱阳湖溯河洄游型刀鲚资源恢复效果初报[J]. 渔业科学进展, 43(1): 24-30. |
姜涛, 周昕期, 刘洪波, 等, 2013. 鄱阳湖刀鲚耳石的两种微化学特征[J]. 水产学报, 37(2): 239-244. |
贺刚, 方春林, 吴斌, 等, 2017. 鄱阳湖刀鲚生殖群体特征及状况分析[J]. 水生态学杂志, 38(3): 83-88. |
袁传宓, 1987. 刀鲚的生殖洄游[J]. 生物学通报, 22(12): 1-3. |
袁传宓, 1988. 长江中下游刀鲚资源和种群组成变动状况及其原因[J]. 动物学杂志, 23(3): 12-15. |
袁传宓, 秦安舲, 1984. 我国近海鲚鱼生态习性及其产量变动状况[J]. 海洋科学, 8(5): 35-37. |
袁传宓, 秦安舲, 刘仁华, 等, 1980. 关于长江中下游及东南沿海各省的鲚属鱼类种下分类的探讨[J]. 南京大学学报(自然科学版), 16(3): 67-82. |
徐跑, 徐刚春, 刘凯, 等, 2016. 长江刀鲚种质资源及人工繁养技术[M]. 北京: 科学出版社, 187-189.
|
郭弘艺, 张旭光, 唐文乔, 等, 2016. 长江靖江段刀鲚捕捞量的时间变化及相关环境因子分析[J]. 长江流域资源与环境, 25(12): 1850-1859. |
郭弘艺, 唐文乔, 2006. 长江口刀鲚矢耳石重量与年龄的关系及其在年龄鉴定中的作用[J]. 水产学报, 30(3): 347-352. |
程起群, 李思发, 2004. 刀鲚和湖鲚种群的形态判别[J]. 海洋科学, 28(11): 39-43. |
詹秉义, 2005. 渔业资源评估[M]. 北京: 中国农业出版社, 18-22.
|
廖伏初, 何望, 黄向荣, 等, 2002. 洞庭湖渔业资源现状及其变化[J]. 水生生物学报, 26(6): 623-627. |
黎雨轩, 何文平, 刘家寿, 等, 2010. 长江口刀鲚耳石年轮确证和年龄与生长研究[J]. 水生生物学报, 34(4): 787-793. |
LIU K, YIN D H, SHU Y L, et al, 2019. Transcriptome and metabolome analyses of Coilia nasus in response to Anisakidae parasite infection[J]. Fish & Shellfish Immunology, 87: 235-242. |
SOKTA L, JIANG T, LIU H B, et al, 2020. Loss of Coilia nasus habitats in Chinese freshwater lakes: an otolith microchemistry assessment[J]. Heliyon, 16(8): e04571. |
WANG M Y, XU P, ZHU Z X, 2020. Regulation of signal transduction in Coilia nasus during migration[J]. Genomics, 112(1): 55-64. DOI:10.1016/j.ygeno.2019.07.021 |
ZHANG N, SONG N, GAO T X, 2016. The complete mitochondrial genome of Coilia nasus (Clupeiformes: engraulidae) from Ariake Sea[J]. Mitochondrial DNA Part A, 27(2): 1518-1519. DOI:10.3109/19401736.2014.953119 |
ZHANG S M, XU Z S, LUO L F, et al, 2023. Genetic diversity and population structure of Coilia nasus revealed by 2b-RAD sequencing[J]. Biology, 12(4): 600. DOI:10.3390/biology12040600 |