 2024, Vol. 55
2024, Vol. 55

红鳍东方鲀(Takifugu rubripes)耐低温性状和生长性状遗传参数评估

http://dx.doi.org/10.11693/hyhz20231100251
中国海洋湖沼学会主办。
中国海洋湖沼学会主办。
文章信息
- 赵海池, 刘志峰, 王新安, 包玉龙, 刘圣聪, 杨明超, 闫鹏飞, 马爱军. 2024.
- ZHAO Hai-Chi, LIU Zhi-Feng, WANG Xin-An, BAO Yu-Long, LIU Sheng-Cong, YANG Ming-Chao, YAN Peng-Fei, MA Ai-Jun. 2024.
- 红鳍东方鲀(Takifugu rubripes)耐低温性状和生长性状遗传参数评估
- EVALUATION OF GENETIC PARAMETERS FOR LOW TEMPERATURE TOLERANT TRAITS AND GROWTH TRAITS OF TAKIFUGU RUBRIPES
- 海洋与湖沼, 55(2): 517-525
- Oceanologia et Limnologia Sinica, 55(2): 517-525.
- http://dx.doi.org/10.11693/hyhz20231100251
文章历史
-
收稿日期:2023-11-21
收修改稿日期:2024-01-15
引用本文
赵海池, 刘志峰, 王新安, 包玉龙, 刘圣聪, 杨明超, 闫鹏飞, 马爱军. 2024. 红鳍东方鲀(Takifugu rubripes)耐低温性状和生长性状遗传参数评估[J]. 海洋与湖沼, 55(2): 517-525

ZHAO Hai-Chi, LIU Zhi-Feng, WANG Xin-An, BAO Yu-Long, LIU Sheng-Cong, YANG Ming-Chao, YAN Peng-Fei, MA Ai-Jun. 2024. EVALUATION OF GENETIC PARAMETERS FOR LOW TEMPERATURE TOLERANT TRAITS AND GROWTH TRAITS OF TAKIFUGU RUBRIPES[J]. Oceanologia et Limnologia Sinica, 55(2): 517-525.
红鳍东方鲀(Takifugu rubripes)耐低温性状和生长性状遗传参数评估
1. 海水养殖生物育种与可持续产出全国重点实验室 中国水产科学研究院黄海水产研究所 中国-东盟海水养殖技术"一带一路"联合实验室(青岛) 山东青岛 266071;
2. 上海海洋大学水产与生命学院 上海 201306;
3. 唐山牧海水产养殖有限公司 河北唐山 063205
2. 上海海洋大学水产与生命学院 上海 201306;
3. 唐山牧海水产养殖有限公司 河北唐山 063205
摘要:采用人工控温的方式, 对构建的31个F1红鳍东方鲀(Takifugu rubripes)全同胞家系开展低温胁迫实验, 获得耐低温性状低温累计存活时间(CDH), 基于混合线性模型分别开展耐低温性状和生长性状遗传参数评估, 对每一性状是否需要考虑共同环境效应所构建的两种模型进行似然比检验。结果显示, 经似然比检验, 最终选用模型A和模型BF进行耐低温和生长性状遗传评估; 耐低温性状CDH遗传力为(0.27±0.08), 属于中等遗传力; 体重BW遗传力为(0.36±0.13), 属于中等遗传力, 体长BL遗传力为(0.14±0.06), 属于低等遗传力, 经检验, 遗传力估计值均达到极显著水平(P < 0.01)。CDH和体重、体长的遗传相关分别为(–0.40±0.22)和(–0.44±0.24), 表型相关分别为(–0.09±0.06)和(–0.16± 0.05), 均为负相关; 体重和体长之间的遗传相关为(0.92±0.05), 表型相关为(0.80±0.02), 呈正相关且结果极显著(P < 0.01)。研究结果表明, 红鳍东方鲀的耐低温性状和生长性状都具有较好的改良潜力, 考虑到两性状间存在负遗传相关, 在开展耐低温选育时, 对首先不同性状进行品系选育, 然后利用品系间杂交培育出耐高温、生长快的新品种。该项研究首次完成了红鳍东方鲀耐低温性状的遗传参数评估, 为制订红鳍东方鲀耐低温选育育种规划提供了理论依据。
关键词:红鳍东方鲀(Takifugu rubripes) 耐低温性状 遗传力 遗传相关
EVALUATION OF GENETIC PARAMETERS FOR LOW TEMPERATURE TOLERANT TRAITS AND GROWTH TRAITS OF TAKIFUGU RUBRIPES
1. National Key Laboratory of Mariculture Biobreeding and Sustainable Goods, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, China-ASEAN Belt and Road Joint Laboratory on Mariculture Technology (Qingdao), Qingdao 266071, China;
2. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China;
3. Tangshan Muhai Aquaculture Co., Ltd., Tangshan 063205, China
2. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China;
3. Tangshan Muhai Aquaculture Co., Ltd., Tangshan 063205, China
Abstract: The culture area of Takifugu rubripes is primarily concentrated in the northern regions of China, where winter water temperatures fall significantly below the species' optimal growth range. Overcoming the challenges associated with indoor wintering, such as low survival rates and increasing costs, is essential to promote sustainable development in the industry. Consequently, there is an urgent need to develop new varieties with improved cold tolerance. In this study, we evaluated the response of 31 F1 full-sib families to low-temperature stress through controlled temperature manipulation. We quantified low temperature tolerant traits using cooling degree hours (CDH) as a trait. To assess the genetic parameters of both low temperature tolerant traits and growth-related traits, we established mixed linear models. The likelihood ratio is used to verify and compare the different models, and finally model A and model BF are selected. Our findings indicate that CDH heritability for low temperature tolerant trait was estimated at 0.27±0.08, reflecting a medium heritability. Heritability values for body weight (BW) as growth trait was 0.36±0.13 indicating a medium heritability. The heritability of body weight was 0.14±0.06, reflecting a low heritability. These estimates reached a highly significant level (P < 0.01) upon testing. Genetic correlations between CDH and both body weight and body length were –0.40±0.22 and –0.44±0.24, respectively, with phenotypic correlations of –0.09±0.06 and –0.16±0.05, all indicating negative associations. In contrast, the genetic correlation between body weight and body length for growth traits was 0.92±0.05, with a phenotypic correlation of 0.80±0.02, demonstrating a strong positive relationship (P < 0.01). Our results suggest that there is significant potential for improving both low temperature tolerant traits and growth traits in T. rubripes. When developing breeding plans, it is crucial to consider trait correlations. One strategy could involve selecting for low-temperature tolerance and fast growth separately and then crossing to create new varieties with composite traits. This study represents the first comprehensive evaluation of genetic parameters related to low-temperature tolerance in T. rubripes, offering valuable theoretical support for the development of new varieties with improved cold tolerance.
Key words:
Takifugu rubripes low temperature tolerant traits heritability genetic correlation
红鳍东方鲀(Takifugu rubripes)是一种重要的海水养殖鱼类, 因其营养丰富, 肉质鲜美, 有“鱼中之王”的美誉。红鳍东方鲀属于暖温性、广温性鱼类, 适宜生长水温为14~27 ℃, 最适水温为16~23 ℃ (马爱军等, 2014)。养殖区域在我国主要集中在辽宁、河北和山东地区, 由于北方冬季的极端低温, 红鳍东方鲀需要进行室内越冬, 不可避免地会面临着成活率降低以及因水温提升带来的成本上升等问题, 因此选育红鳍东方鲀耐低温新品种对扩大良种覆盖率、维持其产业可持续发展具有重要作用。
制定合理的育种规划是品种选育能否成功的关键, 遗传参数是制定育种规划的前提和基础。遗传力作为最重要的一个基本遗传参数, 是数量遗传学中由表及里, 从表型变异研究其遗传实质的一个关键的定量指标。在育种值估计、选择指数指定、选择反应预测、选择方法比较以及育种规划决策等方面, 均起着十分重要的作用(马爱军等, 2008)。遗传力通过量化遗传因素在总体方差中的贡献, 揭示了个体遗传背景在目标性状表现中的重要程度(王松林等, 2023)。通过精确遗传力估计, 不仅能够预测性状在后代中的变异情况, 还能为育种计划的设计和实施提供实际依据(孙坤, 2021)。遗传相关是数量遗传学中另一个重要的基本遗传参数, 可用来描述不同性状之间由于各种遗传原因造成的相关程度大小。在确定间接选择的依据和预测间接选择反应大小、比较不同环境条件下的选择效果以及综合选择指数的制定等方面具有重要的作用(马爱军等, 2008)。在多性状育种中, 通过分析性状间的遗传相关, 可为制订多性状育种规划提供理论依据。一般来说, 抗逆性状和生长性状多呈负相关(蒋湘等, 2014), 因此最好是选择通过定向选育不同的品系再进行杂交的方式进行新品种选育。
温度是决定鱼类生长和代谢的关键环境因素, 国内外已开展部分关于水产动物耐温性遗传参数的研究。刘宝锁等(2011)通过比较两种动物模型发现共同环境系数为0, 估计大菱鲆(Scophthalmus maximus)关于耐高温性状的遗传参数, 获得耐热性(upper thermal tolerance, UTT)遗传力为0.026±0.034。Thodesen等(2013)获得奥利亚罗非鱼(Oreochromis aureus)两个批次耐低温性状的遗传力分别为0.20± 0.04和0.30± 0.31, 属于中等遗传力水平。Charo-Karisa等(2005)开展尼罗罗非鱼(Oreochromis niloticus)幼鱼的耐寒性研究, 得到低温累计存活时间(cooling degree hours, CDH)的遗传力为0.08±0.19, 死亡时温度(temperature at death, TAD)的遗传力为0.09±0.17, 属于低等遗传力。Wang等(2017)研究中国对虾(Fenneropenaeus chinensis)幼虾耐寒性的遗传参数, 其TAD和CDH的遗传力分别为0.265±0.091和0.077±0.058, 属于中低等遗传力。由此可以看出, 耐温性状的遗传力基本处于中低等水平, 不同物种间存在一定的差异。同时耐温性状的表型选择并不一致, 选择合适的表型能提升遗传参数估计的准确性。
本研究通过室内人工降温的方式对31个全同胞家系1 860尾红鳍东方鲀开展低温胁迫实验, 以CDH作为评定耐低温性状的指标, 利用建立的混合线性模型评估红鳍东方鲀的耐低温性状和生长性状遗传力以及它们之间的遗传相关和表型相关, 为红鳍东方鲀耐低温新品种的选育规划提供一定的理论参考。
1 材料与方法 1.1 实验材料红鳍东方鲀亲鱼来自唐山牧海水产养殖有限公司, 挑选出体格健硕、无外伤的亲鱼, 经过一段时间的营养调控和生殖调节, 在同一时期达到性成熟, 用来构建家系后代。2021~2022年, 采用1雌1雄的交配设计方案, 两批次成功构建了31个F1全同胞家系, 各家系分别在不同的养殖池中饲养并标记家系标号, 当家系达到8月龄后用于耐低温测定实验。
1.2 低温胁迫实验低温胁迫实验在高度为1.2 m, 直径为1.5 m的养殖桶中进行, 每6个养殖桶组成一套系统连接一个控温设备用来循环水降温如图 1所示。实验开始前先利用控温设备将水温维持到15 ℃, 从每个家系中随机选取60条红鳍东方鲀放入单独养殖桶中暂养7 d。暂养结束后, 开始降温实验, 从15 ℃开始每1 h下降1 ℃, 在水温降到11 ℃以后改为每2 h降温1 ℃, 到达5 ℃后维持。实验开始后每小时检查一次养殖桶, 收集冷休克鱼体。冷休克状态为鱼体下沉或鱼体出现侧翻轻触后仍不能回正。准确称量鱼体体重、体长, 记录达到冷休克状态的时间、温度等信息, 并剪取尾鳍低温保存。当实验用鱼全部达到冷休克状态后, 低温胁迫实验结束。
 |
| 图 1 循环水降温系统图 Fig. 1 Cooling system diagram of circulating water |
使用Excel软件对红鳍东方鲀的冷休克状态时的温度和时间、个体的CDH值、体长、体重及家系信息等描述性统计数据分别按照家系水平和个体水平进行整理, 使用SPSS软件对描述性统计数据计算、分析并检验家系间差异显著性及个体水平数据是否近似正态分布。使用ASReml4.1.0软件(Butler et al, 2018)对耐低温性状和生长性状的方差组分、遗传力和遗传相关进行分析。
CDH值表示鱼体在低温胁迫下进入冷休克状态前最初温度与每个小时温度之差乘以累计小时数的总和, 计算CDH的初始温度T0为15 ℃。公式为:
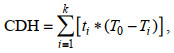 (1)
(1)其中, ti为低温胁迫下的累计小时数, 初始温度度T0为15 ℃, Ti为第i个小时的温度, k为冷休克时的累计小时数。
1.3.2 建立混合线性模型使用ASReml软件建立动物模型, 利用平均信息约束极大似然法(average information restricted maximum likelihood, AIREML) (王明珠等, 2018)计算红鳍东方鲀耐低温和生长性状方差组分。用计算得到的CDH值描述耐低温性状能力, CDH值越大, 表明耐低温能力越强。经检验, 系统Tank做固定效应具有显著性。采用单性状动物模型估计耐低温性状的方差组分, 基于是否考虑共同环境效应构建了两个动物模型, 分别命名为模型A(不含共同环境效应)和模型AF(包含共同环境效应)。
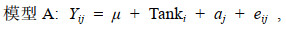 (2)
(2)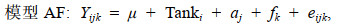 (3)
(3)其中, Yij和Yijk是耐低温性状第j个个体的CDH值, μ是总体均值, Tanki是第i个系统, aj是第j个个体的加性遗传效应, fk是第k个共同环境效应, eij和eijk是随机误差。
估计生长性状遗传力时, 利用单性状动物模型估计生长性状的方差组分, 同样基于是否考虑共同环境效应构建了两个动物模型, 分别命名为模型B(不含共同环境效应)和模型BF(包含共同环境效应)。
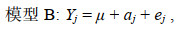 (4)
(4)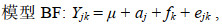 (5)
(5)其中, Yj和Yjk生长性状第j个个体的性状观测值, μ是总体均值, aj是第j个个体的加性遗传效应, fk是第k个共同环境效应, ej和ejk是随机误差。
1.3.3 模型比较利用似然比检验(LRT)比较不同模型, 当比较耐低温性状模型A和模型AF时, 模型A是模型AF的子模型, 当比较生长性状模型B和模型BF时, 模型B是模型BF的子模型。检验统计量如下:
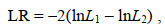 (6)
(6)其中, LR为似然比值, lnL1和lnL2分别是两个不同模型的最大似然函数值, 模型1是模型2的子模型, 统计量符合卡方分布, 自由度为模型2中估计的参数个数与模型1中估计的参数个数之差。
1.3.4 遗传参数的评估表型方差由加性遗传方差、共同环境效应方差和残差组分构成, 计算公式为:
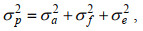 (7)
(7)遗传力的计算公式为:
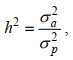 (8)
(8)其中, h2是耐低温或生长性状遗传力, σp2是表型方差, σa2是加性遗传方差, σf2是共同环境效应方差, σe2是残差组分。
在耐低温性状、体重和体长两两之间的遗传和表型相关分析中, 通过两性状动物模型, 估计相应的方差组分和协方差组分, 从而计算得到红鳍东方鲀的耐低温和生长性状的遗传相关和表型相关估计值。
遗传相关、表型相关计算公式为:
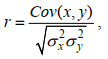 (9)
(9)其中, r是遗传相关或表型相关, Cov(x, y)是遗传协方差组分或表型协方差组分, σx2和σy2是性状的加性遗传方差或表型方差。
1.3.5 显著性检验分析运用Z-score来检验上述获得耐低温和生长性状的遗传力, 以及性状之间遗传和表型相关是否具有显著性(Nguyen et al, 2007):
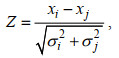 (10)
(10)其中, xi和xj是耐低温和生长性状的遗传力或性状之间的遗传和表型相关, σi和σj是指相对应的标准误。当| Z |≥1.96, 认为遗传力或性状之间的遗传和表型相关存在显著差异(P < 0.05); 当| Z |≥2.58, 认为遗传力或性状之间的遗传和表型相关存在极显著差异(P < 0.01)。
2 结果 2.1 红鳍东方鲀耐低温性状和生长性状的描述性统计 2.1.1 家系水平描述性统计红鳍东方鲀各家系CDH的平均值如图 2a所示, 家系22的平均值最大为1 781.5, 家系9的平均值最小为1 145.5, 各家系的标准差为93.46~579.64, 变异系数为0.05~0.41。经Kruskal-Wallis检验, CDH家系间存在显著差异(P < 0.05)。各家系体重的平均值如图 2b所示, 在批次1中平均值为90.14~187.20, 标准差为22.46~49.55, 变异系数为0.18~0.31; 在批次2中平均值为270.32~431.30, 标准差为22.68~109.77, 变异系数为0.07~0.29。各家系体长的平均值如图 2c所示, 在批次1中体长平均值为13.68~16.54, 标准差为0.96~1.69, 变异系数为0.06~ 0.11; 在批次2中体长平均值为20.06~22.50, 标准差为0.54~1.81, 变异系数为0.03~0.08。经Kruskal-Wallis检验, 体重和体长分别在两个批次家系间存在显著差异(P < 0.05)。以上结果见表 1。
 |
| 图 2 红鳍东方鲀CDH、体重和体长家系水平的平均值直方图 Fig. 2 Histogram of the average value of CDH, body weight and body length at family level to T. rubripes |
表 1 红鳍东方鲀耐低温性状和生长性状家系水平的描述性统计 Tab. 1 The descriptive statistics of low temperature tolerant traits and growth traits at family level to T. rubripes
| 性状 | 批次 | 样本数 | 平均值 | 标准差(SD) | 变异系数CV/% |
| CDH | — | 31 | 1 145.5~1 781.5 | 93.46~579.64 | 0.05~0.41 |
| 体重BW/g | 1 | 13 | 90.14~187.20 | 22.46~49.55 | 0.18~0.31 |
| 2 | 18 | 270.32~431.30 | 22.68~109.77 | 0.07~0.29 | |
| 体长BL/cm | 1 | 13 | 13.68~16.54 | 0.96~1.69 | 0.06~0.11 |
| 2 | 18 | 20.06~22.50 | 0.54~1.81 | 0.03~0.08 |
耐低温性状CDH、体重和体长的样本数、平均值、标准差和变异系数等描述性统计数据如表 2所示, CDH、体重和体长的平均值分别为1 459、230.42 g、18.11 cm。体重的范围为22.49~586.30 g, 变异系数较大(CV= 47.90%)。体长的标准差为3.18, 具有较小的变异系数(CV=17.54%)。耐低温性状指标CDH有较大的变异系数(CV=28.68%), 表明红鳍东方鲀的个体间耐低温能力有较大的差异, 耐低温性状具有较好的改良潜力。
表 2 红鳍东方鲀耐低温性状和生长性状的描述性统计 Tab. 2 The descriptive statistics of low temperature tolerant traits and growth traits to T. rubripes
| 性状 | 样本数 | 平均值 | 最小值 | 最大值 | 标准差SD | 变异系数CV/% |
| CDH | 1 860 | 1 459 | 380 | 1 965 | 418.3 | 28.68 |
| 体重BW/g | 1 860 | 230.42 | 22.49 | 586.30 | 110.37 | 47.90 |
| 体长BL/cm | 1 860 | 18.11 | 11.10 | 25.20 | 3.18 | 17.54 |
红鳍东方鲀个体水平CDH、体重和体长数据分别拟合正态分布如图 3所示, CDH的拟合度(R2)为0.799 57, 其余生长性状数据的拟合度较高都大于0.95, 其中体重两个批次拟合度分别为0.991 53和0.985 75, 体长两个批次的拟合度分别为0.992 7和0.997 96, 通过方差分析表明数据近似正态分布具有统计学意义(P < 0.05)。
 |
| 图 3 红鳍东方鲀CDH、体重和体长频率分布直方图 Fig. 3 Frequency distribution histogram of CDH, body weight and body length to T. rubripes |
利用似然比验证(LRT)比较不同模型, 分析耐低温性状和生长性状模型中是否需要考虑共同环境效应结果见表 3。耐低温性状两个模型比较, 模型A与模型AF的似然值相同两个模型之间无显著差异(P > 0.05)。体重性状两个模型比较, 模型B与模型BF的似然值不相同, 似然比为82.223, 两个模型差异极显著(P < 0.01)。体长性状两模型比较, 模型B与模型BF的似然值也不相同, 似然比为137.52, 两个模型差异极显著(P < 0.01)。因此, 估计耐低温性状遗传参数选用不考虑共同环境效应的模型A, 估计生长性状遗传参数则选用加入共同环境效应的模型BF。
表 3 不同动物模型似然值及似然比 Tab. 3 Likelihood value and likelihood ratio of different animal models
| 性状 | 模型 | 似然值(–2 ln) | 似然比(LR) |
| CDH | A | 9 564.479 | |
| AF | 9 564.479 | 0 | |
| 体重BW | B | 6 857.578 | |
| BF | 6 779.984 | 82.223** | |
| 体长BL | B | 1 396.187 | |
| BF | 1 163.295 | 137.52** | |
| 注: **表示结果达到极显著水平(P < 0.01), 下同 | |||
红鳍东方鲀耐低温和生长性状的方差组分和遗传力如表 4所示, 耐低温性状CDH遗传力为0.27± 0.08, 属于中等遗传力, 体重遗传力为0.36±0.13, 属于中等遗传力, 体长遗传力为0.14±0.06, 属于低遗传力。经检验, 所获得的遗传力均极显著(P < 0.01)。
表 4 红鳍东方鲀耐低温和生长性状方差组分和遗传力 Tab. 4 Variance components and heritability of low temperature tolerant traits and growth traits to T. rubripes
| 性状 | 方差组分 | 遗传力h2 | |||
| 加性遗传方差σa2 | 共同环境效应σf2 | 残差组分σe2 | 表型方差σp2 | ||
| CDH | 34 273.89 | — | 89 881.45 | 124 155.34 | 0.27±0.08** |
| 体重BW | 2 660.43 | 3 506.84 | 1 110.60 | 7 277.87 | 0.36±0.13** |
| 体长BL | 0.67 | 3.05 | 1.15 | 4.87 | 0.14±0.06** |
红鳍东方鲀的耐低温性状CDH、体重和体长两两之间估计的遗传相关和表型相关结果如表 5所示。用CDH来表示耐低温性状指标, CDH与体重和体长的遗传相关分别为–0.40±0.22、–0.44±0.24, 表型相关分别为–0.09±0.06、–0.16±0.05 (P < 0.01), 均为负相关。体重和体长两者间的遗传和表型相关分别为0.92±0.05、0.80±0.02, 生长性状的两个指标体重与体长之间呈高度正相关(P < 0.01)。
表 5 红鳍东方鲀CDH、体重和体长之间的遗传表型相关性 Tab. 5 Genetic and phenotypic correlation between CDH, body weight and body length to T. rubripes
| 性状 | CDH | 体重BW | 体长BL |
| CDH | — | –0.40±0.22 | –0.44±0.24 |
| 体重BW | –0.09±0.06 | — | 0.92±0.05** |
| 体长BL | –0.16±0.05** | 0.80±0.02** | — |
| 注: 上三角为遗传相关, 下三角为表型相关 | |||
水产动物耐低温选育是近年来的热点领域, 关于耐低温的研究已开展了大量的报道, 包括低温胁迫下的生理反应(Liu et al, 2022)、相关基因表达(刘志峰等, 2023)、分子标记筛选(袁晨浩等, 2022)等。然而, 关于水产动物耐低温性状的遗传力报道少有研究, 目前仅罗非鱼(Charo-Karisa et al, 2005; Thodesen et al, 2013)、中国对虾(Li et al, 2016; Wang et al, 2017)、凡纳滨对虾(Penaeus vannamei) (Li et al, 2015)等有耐低温性状遗传力的报道, 还没有关于红鳍东方鲀耐低温性状遗传评估的研究。研究人员通常会选择CDH、TAD、半致死时间的存活状态(Survival status at half lethal time, SS50)等作为描述耐低温能力的指标, 如卢其西等(2011)用CDH做为奥利亚罗非鱼的耐寒指标, 评价6个家系的耐寒能力, 王明珠等(2018)和Wang等(2017)利用TAD、CDH和SS50分别描述中国对虾及其幼虾的耐低温程度。一般来说, CDH结合了低温存活的持续时间和温度, 描述耐低温性状更精确, 因此本研究使用CDH作为指标来评估红鳍东方鲀的耐低温能力并进行遗传参数评估。
本研究获得CDH的遗传力为0.27±0.08, 与奥利亚罗非鱼(Thodesen et al, 2013)、中国对虾(王明珠等, 2018)研究得到的耐低温性状遗传力水平相似, 都属于中等遗传力水平。然而, 这个结果并不是普遍现象, Li等(2015, 2016)研究了凡纳滨对虾和中国对虾的耐寒性遗传力, 凡纳滨对虾的SS50遗传力为0.021 1±0.019 6, CDH遗传力为0.025 8±0.020 5; 中国对虾SS50遗传力为0.059 3±0.016 0, 加入共同环境效应后遗传力下降为0.019 2±0.023 5。Charo-Karisa等(2005)用CDH和TAD两种指标来描述耐低温性状, 估计尼罗罗非鱼幼鱼耐低温性状的遗传力, 获得CDH和TAD的遗传力分别为0.08±0.17和0.09±0.19。这些遗传力结果都是低遗传力, 明显低于本研究的结果。我们推测这种差异主要来源于种间差异, 此外, 遗传评估的个体数量、不同的统计方法和模型、以及所用样本的不同发育时期同样是不可忽视的因素。
3.2 红鳍东方鲀生长性状遗传力生长性状作为重要的经济性状, 近几年已开展大量关于水产动物生长性状遗传参数的研究, 如菲律宾蛤仔(Ruditapes philippinarum) (王松林等, 2023)、全州禾花鲤(Cyprinus carpio var. Quanzhouensis) (许艺兰等, 2022)、金鲷(Chrysophrys auratus) (Ashton et al, 2019)和凡纳滨对虾(王伦等, 2022)等。针对红鳍东方鲀生长性状的遗传力估计也有部分研究, 刘永新等(2014)以135日龄生长性状为协变量, 构建了三个不同的动物模型用来估计红鳍东方鲀200日龄的生长性状遗传力。通过比较不同模型, 确定有全同胞效应并且不含母本效应的模型三为最优模型, 所估计的体质量和体长遗传力分别为0.16和0.14。刘永新等(2017)又估计红鳍东方鲀150日龄和240日龄的体质量遗传力分别为0.17和0.21, 体长遗传力分别为0.15和0.18。两次对红鳍东方鲀的生长性状估计所选择的群体日龄接近, 并且结果相近都属于中低等遗传力。这与本研究的结果相似, 本研究的结果发现体重的遗传力为0.36±0.13, 属于中等遗传力, 体长的遗传力为0.14±0.06, 属于低等遗传力。于海龙等(2016)基于仅加性遗传效应作随机效应的动物模型, 估计红鳍东方鲀210日龄和360日龄生长性状遗传力, 获得红鳍东方鲀体质量遗传力为0.54和0.46, 体长遗传力为0.38和0.37, 全长遗传力为0.52和0.44, 这与本研究的结果并不相同, 属于中高等遗传力。分析原因可能是忽略了共同环境效应, 没有将共同环境效应从加性遗传效应中准确地剖分出来, 导致结果偏大。马文超等(2021)为了探究红鳍东方鲀在不同生长时期的生长性状遗传力, 分别估计了1、2、6和18月龄的体重、体长和全长的遗传力, 得到各个生长时期的体重遗传力分别为0.73、0.48、0.23和0.23, 体长遗传力分别为0.65、0.38、0.19和0.16、全长遗传力分别为0.66、035、0.16和0.19。其中, 6月龄与本研究使用的实验鱼月龄接近, 得到该月龄的结果属于中低等遗传力, 与本研究得到的结果一致。此外, 不难看出随着月龄的增长生长性状遗传力呈下降趋势, 因所选用的方法无法考虑母本效应的影响, 并且母本效应在生物发育早期占有一定的方差组分(Shimada et al, 2007), 因此1月龄和2月龄的生长性状遗传力估计值偏大。红鳍东方鲀的生长性状遗传力估计值存在差异, 主要是由于共同环境效应和母本效应是否考虑加入到模型中, 不同的动物模型估计的遗传力往往差异较大(王延晖等, 2023), 选择合适的模型能提高遗传力估计的准确性。
3.3 红鳍东方鲀耐低温性状和生长性状之间的相关分析除了遗传力之外, 遗传相关也是重要的遗传参数。当两个性状之间的相关性较高时, 对其中一个性状的选择育种也会对另一个性状有较大的影响。因此对于那些改良难度大或是难以直接测量的性状, 可以利用与该性状相关性强的其他性状进行间接选择, 当目标性状间有较强的负相关时, 由于性状间存在相互拮抗, 需要改变育种策略, 首先对不同性状进行品系选育, 然后利用品系间杂交培育出不同性状同时得到改良的新品种。本研究中红鳍东方鲀CDH与体重的遗传相关和表型相关分别为–0.40±0.22和–0.09±0.06, 与体长的遗传相关和表型相关分别为–0.44±0.24和–0.16±0.05, 均为负相关, 且遗传相关高于表型相关。这与其他研究中的结果相类似, 比如Li等(2015)对凡纳滨对虾的耐低温遗传参数做了研究, CDH和SS50与体重之间存在较强的负相关, 遗传相关分别为–0.770 2±0.458 3和–0.825 3±0.455 3, 表型相关分别为0.028 4±0.027 6和–0.062 8±0.201 1。Charo-Karisa等(2005)研究尼罗罗非鱼幼鱼耐低温性状发现体重与耐低温性状TAD的遗传相关和表型相关分别为–0.68±0.84和–0.26±0.18, 为负相关。另外有些研究结果也表明, 耐低温性状与生长性状之间存在一定的正相关, 王明珠等(2018)研究中国对虾耐低温性状与生长性状的相关性, 体重与SS50的遗传相关和表型相关分别为0.517±0.205和0.187±0.030, 体长与SS50的遗传相关和表型相关分别为(0.538±0.203)和(0.218±0.030)。由此可以看出, 不同的物种, 其抗逆性状与生长表型的相关性并不一定是普遍的负相关, 这对育种的路线规划至关重要, 也从侧面反映出遗传参数评估的重要性。
4 结论本研究利用构建的31个F1全同胞家系, 开展红鳍东方鲀耐低温性状和生长性状的遗传参数评估。获得耐低温性状遗传力为中等遗传力, 生长性状体重遗传力为中等遗传力, 体长遗传力为低等遗传力, 表明通过群体选育或者家系选育对红鳍东方鲀耐低温性状和生长性状进行改良, 能够获得较好的遗传进展。另外, 耐低温性状和生长性状的表型相关和遗传相关为负相关, 由于性状间存在相互拮抗, 因此制定育种方案时, 首先对不同性状进行品系选育, 然后利用品系间杂交培育出不同性状同时得到改良的新品种。研究结果为红鳍东方鲀耐低温新品种的选育规划提供一定的理论参考。
参考文献
于海龙, 顾晨雷, 孔德荣, 等, 2016. 红鳍东方鲀生长性状的遗传力估计及性状间相关关系的初步研究[J]. 大连海洋大学学报, 31(4): 357-361. |
马文超, 杨晓, 王宁, 等, 2021. 红鳍东方鲀不同阶段生长性状的遗传力估计[J]. 浙江海洋大学学报(自然科学版), 40(5): 407-414, 448. |
马爱军, 王新安, 杨志, 等, 2008. 大菱鲆(Scophthalmus maximus)幼鱼生长性状的遗传力及其相关性分析[J]. 海洋与湖沼, 39(5): 499-504. |
马爱军, 王新安, 雷霁霖, 2009. 大菱鲆(Scophthalmus maximus)不同生长阶段体重的遗传参数和育种值估计[J]. 海洋与湖沼, 40(2): 187-194. |
马爱军, 李伟业, 王新安, 等, 2014. 红鳍东方鲀养殖技术研究现状及展望[J]. 海洋科学, 38(2): 116-121. |
王延晖, 王冰柯, 屈长义, 等, 2023. 不同模型估计黄河鲤生长性状遗传参数的比较分析[J]. 水产科学, 42(3): 377-385. |
王伦, 王崇懿, 刘建勇, 2022. 凡纳滨对虾生长和耐综合胁迫性状的遗传参数评估[J]. 南方水产科学, 18(4): 95-102. |
王松林, 孙秀俊, 涂康, 等, 2023. 菲律宾蛤仔早期生长性状的遗传参数评估及生长模型构建[J]. 中国海洋大学学报, 53(9): 69-76. |
王明珠, 孟宪红, 孔杰, 等, 2018. 低温胁迫条件下中国明对虾生长性状和耐低温性状的遗传参数评估[J]. 渔业科学进展, 39(3): 96-102. |
卢其西, 林勇, 宾石玉, 等, 2011. 罗非鱼6个家系的低温耐寒测定分析[J]. 广西师范大学学报: 自然科学版, 29(2): 104-109. |
刘永新, 周勤, 张红涛, 等, 2014. 红鳍东方鲀(Takifugu rubripes)生长性状的遗传参数估计[J]. 渔业科学进展, 35(6): 39-44. |
刘永新, 周勤, 张红涛, 等, 2017. 基于标记系谱的红鳍东方鲀生长性状遗传分析[J]. 水产学报, 41(1): 21-30. |
刘志峰, 马爱军, 孙建华, 等, 2023. 红鳍东方鲀AFP、CIRP、HMGB1和YB-1基因对低温胁迫的响应[J]. 渔业科学进展, 44(2): 58-67. |
刘宝锁, 张天时, 孔杰, 等, 2011. 大菱鲆生长和耐高温性状的遗传参数估计[J]. 水产学报, 35(11): 1601-1606. |
许艺兰, 陈忠, 覃俊奇, 等, 2022. 全州禾花鲤主要生长性状的遗传参数估计[J]. 广西科学, 29(4): 801-808. |
孙坤, 2021. 凡纳滨对虾重要经济性状的遗传评估及全基因组关联分析[D]. 上海: 上海海洋大学: 5.
|
袁晨浩, 刘志峰, 马爱军, 2022. 红鳍东方鲀(Takifugu rubripes)耐低温相关微卫星标记的初步筛选[J]. 海洋科学, 46(2): 97-104. |
蒋湘, 刘建勇, 赖志服, 2014. 九孔鲍(Haliotis diversicolor supertexta)耐低盐与生长性状的遗传参数评估[J]. 海洋与湖沼, 45(3): 542-547. |
ASHTON D T, HILARIO E, JAKSONS P, et al, 2019. Genetic diversity and heritability of economically important traits in captive Australasian snapper (Chrysophrys auratus)[J]. Aquaculture, 505: 190-198. DOI:10.1016/j.aquaculture.2019.02.034 |
BUTLER D G, CULLIS B R, GILMOUR A R, et al, 2018. ASReml-R Reference Manual Version 4[M]. Hemel Hempstead, UK: VSN International Ltd.
|
CHARO-KARISA H, REZK M A, BOVENHUIS H, et al, 2005. Heritability of cold tolerance in Nile tilapia, Oreochromis niloticus, juveniles[J]. Aquaculture, 249(1/2/3/4): 115-123. |
LI W J, LUAN S, LUO K, et al, 2015. Genetic parameters and genotype by environment interaction for cold tolerance, body weight and survival of the Pacific white shrimp Penaeus vannamei at different temperatures[J]. Aquaculture, 441: 8-15. DOI:10.1016/j.aquaculture.2015.02.013 |
LI W J, LUAN S, LUO K, et al, 2016. Genetic parameters for cold tolerance and body weight of Chinese fleshy prawn, Fenneropenaeus chinensis[J]. Acta Oceanologica Sinica, 35(6): 12-18. DOI:10.1007/s13131-016-0845-6 |
LIU Z F, MA A J, YUAN C H, et al, 2022. Kidney transcriptome analysis of tiger puffer (Takifugu rubripes) uncovers key pathways and genes in response to low-temperature stress[J]. Aquaculture Reports, 26: 101287. DOI:10.1016/j.aqrep.2022.101287 |
NGUYEN N H, KHAW H L, PONZONI R W, et al, 2007. Can sexual dimorphism and body shape be altered in Nile tilapia (Oreochromis niloticus) by genetic means?[J]. Aquaculture, 272(Suppl 1): S38-S46. |
SHIMADA Y, SHIKANO T, MURAKAMI N, et al, 2007. Maternal and genetic effects on individual variation during early development in Japanese flounder Paralichthys olivaceus[J]. Fisheries Science, 73(2): 244-249. DOI:10.1111/j.1444-2906.2007.01330.x |
THODESEN J, RYE M, WANG Y X, et al, 2013. Genetic improvement of tilapias in China: genetic parameters and selection responses in growth, pond survival and cold-water tolerance of blue tilapia (Oreochromis aureus) after four generations of multi-trait selection[J]. Aquaculture, 396/399: 32-42. DOI:10.1016/j.aquaculture.2013.02.010 |
WANG M Z, KONG J, MENG X H, et al, 2017. Evaluation of genetic parameters for growth and cold tolerance traits in Fenneropenaeus chinensis juveniles[J]. PLoS One, 12(8): e0183801. DOI:10.1371/journal.pone.0183801 |