 2016, Vol. 47
2016, Vol. 47中国海洋湖沼学会主办。
文章信息
- 杜美荣, 方建光, 包振民, 高亚平, 房景辉, 蒋增杰. 2016.
- DU Mei-Rong, FANG Jian-Guang, BAO Zhen-Min, GAO Ya-Ping, FANG Jing-Hui, JIANG Zeng-Jie. 2016.
- 不同养殖海域栉孔扇贝(Chlamys farreri)混合家系的通径分析
- CORRELATION AND PATH COFFICIENT ANALYSIS FOR CHLAMYS FARRERI CULTURED IN DIFFERENT SEA AREAS
- 海洋与湖沼, 47(5): 963-970
- Oceanologia et Limnologia Sinica, 47(5): 963-970.
- http://dx.doi.org/10.11693/hyhz20160100011
-
文章历史
- 收稿日期:2016-01-18
- 收修改稿日期:2016-05-19




2. 农业部海洋渔业资源可持续利用重点开放实验室 山东省渔业资源与生态环境重点实验室 中国水产科学研究院黄海水产研究所 青岛 266071;
3. 青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266000
2. Key Laboratory for Sustainable Development of Marine Fisheries, Ministry of Agriculture, Shandong Provincial Key Laboratory of Fishery Resources and Eco-Environment, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Science, Qingdao 266071, China;
3. Function Laboratory for Marine Fisheries Science and Food Production Processes, Qingdao National Laboratory for Marine Science and Technology, Qingdao 266000, China
栉孔扇贝(Chlamys farreri)分布于我国的山东、辽宁等沿海海域, 因其氨基酸种类和含量丰富、味道鲜美, 是我国北方主要的浅海浮筏养殖品种(王如才等, 1989; 魏利平, 2000), 其产量在90年代几乎达到全国扇贝总产量的80%(Guo et al, 1999)。我国北方养殖的三大扇贝种类, 即栉孔扇贝、海湾扇贝(Argopecten irradias)和虾夷扇贝(Patinopecten yessoensis)中, 只有栉孔扇贝是我国的土著种类。随着20世纪50年代开展栉孔扇贝繁殖学研究获得成功后(张玺等, 1956), 70年代开展了大规模室内苗种的人工培育(王子臣, 1977; 王如才等, 1986), 并先后在大连、青岛和烟台等地培育成功。80年代末期育苗技术愈加成熟, 使得我国的栉孔扇贝人工育苗达到一个鼎盛时期, 养殖用苗种主要来源于人工培育的苗种(魏利平等, 1985; 于瑞海等, 1992)。从1996年开始, 栉孔扇贝在养殖过程中出现大面积死亡现象, 产业遭受重创(张福绥等, 1999a)。多位学者从育苗、养殖和病害等诸方面开展了研究(宋微波等, 2001; 王秀华等, 2002; 于瑞海等, 2006; 王玲玲等, 2007; 杜美荣等, 2009)。有学者认为上世纪90年代栉孔扇贝的大规模死亡与种质的累代使用及种质退化有一定关系(张福绥等, 1999b)。种质的筛选对养殖生物尤为重要, 使用性状优良的亲本培育出的子代才具有良好的性状。体重性状是影响产量的重要指标之一, 也是选育优良品种最直接的经济性状。扇贝体重受多基因控制, 同时又受到其它壳尺寸等数量性状的影响, 但体重性状不直观, 壳尺寸性状则相对比较直观可见。因此使用大量扇贝数据的通径分析可以获得壳尺寸性状与体重性状之间的多元回归分析, 通过对直观的壳尺寸性状选择达到选育种的目的, 指导种贝挑选工作, 具有极其重要的现实意义。
通径分析理论由数量遗传学家Wright(1984)在20世纪20年代提出, 它可以确定参数间的相关性, 并将其相关系数拆分为直接影响和通过其它参数的间接影响, 从而建立最优回归方程。使用通径分析指导水产生物的选育种工作在白斑红点鲑(Salvelinus leucomaenis)、天津厚蟹(Helice tientsinensis)、日本囊对虾(Penaeus japonicus)和日本沼虾(Macrobrachium nipponensis)以及其它多种贝类中都有应用(高玮玮等, 2009; 孙振兴等, 2010; 王志铮等, 2011; 平洪领等, 2013; 张永泉等, 2014; 李洪鹏等, 2015)。影响几种贝类湿重的主要形态性状因子各不相同:针对日本镜蛤(Dosinia japonica Reeve)的研究表明, 影响其体重的主要性状是壳高(张伟杰等, 2013); 壳长对砂海螂(Mya arenaria)的总体重和软体部重的直接效应均最大(孙振兴等, 2013); 针对作为珍珠的主要插核原料背瘤丽蚌(Lamprotula leai)的研究表明, 影响壳重的主要因素为总湿重(张根芳等, 2014); 影响企鹅珍珠贝(Pteria penguin)湿重的主要因素为壳长(粟志民等, 2011)。
生物的表现型受基因型、环境、以及基因型与环境互作几个方面的影响。简而言之, 基因型的表达会随环境的不同而不同(黄英姿等, 1992)。环境以及基因型与环境互作(G×E互作)的存在, 使得同样一组家系在不同环境中的表型性状表现出不同的次序, 即在一个环境中表现好的基因型在另一个环境中并不一定会表现好。虽有针对不同性别以及苗种来源的栉孔扇贝的通径分析(刘小林等, 2002; 杜美荣等, 2012; 王冲, 2013), 但大多数材料的遗传背景尚不清晰, 且养殖海域不同, 养殖环境和饵料背景不清晰, 材料缺乏说服力。本研究以2013年春季建立的11(雌)×10(雄)栉孔扇贝混合家系为材料, 在三个不同的养殖纬度条件下进行养殖实验, 通过测量其生长性状, 研究基于相同遗传背景的栉孔扇贝在不同地理纬度的养殖海区养殖一年后壳尺寸性状对总体重的通径分析差异, 建立壳尺寸性状和总体重性状间的回归方程, 以期建立不同地理环境栉孔扇贝选育种工作中种贝挑选的策略和方案, 为栉孔扇贝选育种工作提供理论支持。
1 材料和方法 1.1 实验材料于2013年4月采用室内升温育苗的方法构建了11(雌)×10(雄)栉孔扇贝混合家系子代个体, 贝苗完成室内培育后, 装进保苗袋于青岛流清河扇贝养殖区进行中间培育, 于当年秋季移入暂养笼进行养成管理。2014年4月, 在混合家系子代12月龄时, 随机选取976个个体对壳长、壳高、壳宽和总体重进行测量并分别标记后分成三份, 分别养殖于不同地理纬度的养殖环境下, 养殖地点分别为青岛流清河扇贝养殖区(36°06′N, 120°32′E)(简称青岛), 荣成桑沟湾扇贝养殖区(37°18′N, 122°45′E)(简称荣成), 大连獐子岛扇贝养殖区(39°03′N, 124°47′E)(简称大连)。经过一周年的养殖, 于2015年4月收回所有扇贝进行壳长、壳高、壳宽和总体重的测定。
1.2 实验方法 1.2.1 数据测定对上述栉孔扇贝数据进行形态学测定, 使用精确度为0.01mm的游标卡尺测定壳长(x1), 壳高(x2), 壳宽(x3), 使用精确度为0.01的天平称量总体重(y)。
1.2.2 数据分析采用SPSS Statistics17.0对数据进行分析。采用One-way ANOVA计算方差, 并用Duncan法进行多重比较, 对总体重进行通径分析, 计算决定系数。数量性状统计数据采用平均值±标准差(M±SD)表示。
对所有表型性状进行相关性分析以及壳尺寸性状对总体重的通径分析, 相关系数rij减去通径系数Pi获得间接通径系数总和Σ, 剖分间接通径系数获得各个性状的间接通径系数rijPj。根据以下公式计算单参数决定系数(di)和多参数决定系数(dij)(孙振兴等, 2010):
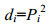 (1)
(1)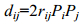 (2)
(2)(1)式中, Pi为某性状对总体重的通径系数; (2)式中, rij为性状间的相关系数, Pi、Pj为相应的壳尺寸性状对总体重的通径系数。
多元线性回归方程为:
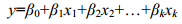 (3)
(3)(3)式中, β0为常数项; β1、β2、…、βk为偏回归系数; x1、x2、…、xk为偏回归系数所对应的自变量。
2 结果 2.1 壳尺寸性状与总体重的统计所得壳长、壳宽和壳高数据见表 1。从三个养殖海域栉孔扇贝的壳尺寸来看:大连海域的壳长最大, 青岛海域的次之; 壳高却是青岛海域的最大, 大连海域的次之; 大连海域栉孔扇贝的壳长与壳高比值最大, 青岛海域的最小。由湿重比较可见, 青岛海域的最大, 大连海域的次之, 荣成海域的稍小(差异显著性比较结果见表 1)。由总体重比较可见, 青岛海域的最大, 大连海域的次之, 荣成海域的稍小。结合壳尺寸性状和总体重性状可以看出, 大连海域栉孔扇贝的壳长和壳宽最大, 但总体重却稍逊于青岛海域的栉孔扇贝, 而荣成海域栉孔扇贝在壳尺寸和总体重方面均不占优势。以上结果表明栉孔扇贝壳尺寸性状具有较强的环境可塑性。由变异系数可知, 总体重的变异系数大于其它壳尺寸性状, 其中大连海域栉孔扇贝总体重变异系数最大, 荣成海域的次之, 青岛海域的最小。变异系数的大小是进行选育的关键参考依据, 变异系数较大时选择的潜力也越大, 开展选择育种的价值也越高。
| 养殖海域 | 参数 | 壳长 | 壳高 | 壳宽 | 总体重 | 长高比 |
| 青岛(n=240) | 平均值 | 65.66 a | 71.63a | 23.79a | 47.38a | 0.92a |
| 标准差 | 6.52 | 5.44 | 1.93 | 8.76 | ||
| 变异系数/% | 9.93 | 7.60 | 8.12 | 18.49 | ||
| 荣成(n=227) | 平均值 | 62.80b | 67.59b | 21.12b | 35.45b | 0.93b |
| 标准差 | 4.65 | 5.03 | 1.81 | 6.61 | ||
| 变异系数/% | 7.40 | 7.44 | 8.55 | 18.63 | ||
| 大连(n=230) | 平均值 | 66.53a | 70.45a | 24.04a | 46.04a | 0.94b |
| 标准差 | 7.01 | 6.98 | 2.78 | 13.46 | ||
| 变异系数/% | 10.54 | 9.90 | 11.56 | 29.24 | ||
| 注:表中壳长、壳宽、壳高的数据单位为mm, 总体重为g; 数据上标的a、b表示数据间的差异显著性 | ||||||
栉孔扇贝各性状间的相关性分析见表 2。本实验中各性状间均表现为极显著相关关系(P < 0.01), 说明壳长、壳高、壳宽与总体重间存在极显著的共线性关系, 使用壳尺寸性状对总体重进行回归分析具有实际意义。其中荣成海域、大连海域与其总体重相关系数最大的性状是壳长, 其次是壳高, 壳宽最小。与青岛海域栉孔扇贝总体重相关系数最大的为壳高, 其次是壳宽, 壳高最小。单一性状与总体重间的相关系数表示该性状对总体重的直接作用以及其它性状通过该性状所起到的间接作用, 因而单纯的相关系数不能确定变量的重要性, 还需用通径分析揭示其它性状对总体重的影响效应, 因此有必要对这种相关关系作进一步分解。
| 养殖海域 | 参数 | 总体重 | 壳长 | 壳高 | 壳宽 |
| 青岛 | 总体重 | 1.00 | 0.69** | 0.81** | 0.77** |
| 壳长 | — | 1.00 | 0.66 ** | 0.48** | |
| 壳高 | — | — | 1.00 | 0.56** | |
| 壳宽 | — | — | — | 1.00 | |
| 荣成 | 总体重 | 1 | 0.91** | 0.81** | 0.79** |
| 壳长 | — | 1.00 | 0.82** | 0.72** | |
| 壳高 | — | — | 1.00 | 0.62** | |
| 壳宽 | — | — | — | 1.00 | |
| 大连 | 总体重 | 1 | 0.89** | 0.88** | 0.87** |
| 壳长 | — | 1.00 | 0.95** | 0.79** | |
| 壳高 | — | — | 1.00 | 0.76** | |
| 壳宽 | — | — | — | 1.00 | |
| 注:表中壳长、壳宽、壳高的数据单位为mm, 总体重为g; *表示P < 0.05, 差异显著, **表示P < 0.01, 差异极显著 | |||||
壳尺寸性状对总体重的通径分析结果如表 3所示。其中对青岛海域栉孔扇贝总体重直接作用最大的是壳高(Pi=0.446), 其次是壳宽(Pi=0.432), 壳长最小(Pi=0.190)。间接作用中, 壳高通过壳长间接影响总体重的程度最大, 其次是壳高通过壳宽, 壳宽通过壳高间接影响总体重的程度最小。
| 养殖海域 | 性状 | 相关系数rij | 直接作用Pi | 间接作用rijPj | |||
| 合计 | 壳长 | 壳宽 | 壳高 | ||||
| 青岛 | 壳长 | 0.690 | 0.190 | 0.500 | — | 0.210 | 0.293 |
| 壳宽 | 0.770 | 0.432 | 0.338 | 0.092 | — | 0.250 | |
| 壳高 | 0.810 | 0.446 | 0.364 | 0.125 | 0.243 | — | |
| 荣成 | 壳长 | 0.913 | 0.598 | 0.315 | — | 0.198 | 0.117 |
| 壳宽 | 0.794 | 0.277 | 0.517 | 0.428 | — | 0.089 | |
| 壳高 | 0.805 | 0.142 | 0.663 | 0.490 | 0.173 | — | |
| 大连 | 壳长 | 0.886 | 0.148 | 0.738 | — | 0.345 | 0.392 |
| 壳宽 | 0.865 | 0.434 | 0.431 | 0.118 | — | 0.313 | |
| 壳高 | 0.883 | 0.413 | 0.470 | 0.141 | 0.329 | — | |
| 注: rij为性状间的相关系数, Pi、Pj为单性状分别对总体重的通径系数 | |||||||
对荣成海域栉孔扇贝总体重直接作用从大到小依次是壳长(Pi=0.598)、壳宽(Pi=0.277)和壳高(Pi=0.142)。壳长通过壳高间接影响总体重的程度最大, 其次是壳长通过壳宽, 壳宽通过壳长间接影响总体重的程度最小。
对大连海域栉孔扇贝总体重直接作用从大到小依次是壳宽(Pi=0.434)、壳高(Pi=0.413)和壳长(Pi=0.148)。壳高通过壳长间接影响总体重的程度最大, 其次是壳宽通过壳长, 壳宽通过壳高间接影响总体重的程度最小。
2.4 壳长、壳高和壳宽对总体重的决定程度分析壳尺寸性状对总体重的决定程度分析结果见表 4, 对角线方向是单参数决定系数, 对角线以上是一个性状通过其它性状间接对总体重的决定系数。青岛海域、荣成海域和大连海域栉孔扇贝各壳尺寸性状对总体重的决定系数总和分别为0.830、0.880和0.872, 与回归方程的R2相等, 说明所选取的壳长、壳高和壳宽都是影响总体重的主要性状。对荣成海域栉孔扇贝总体重直接影响程度最大的是壳长(di=0.358), 对青岛海域栉孔扇贝总体重直接影响程度最大的是壳高(di=0.199), 对大连海域栉孔扇贝总体重直接影响程度最大的是壳宽(di=0.189)。可见, 即便是遗传背景相同的扇贝, 在不同的养殖海域养殖一年后, 对其总体重起决定作用的壳尺寸性状亦有差别。就间接决定系数而言, 对青岛海域和大连海域栉孔扇贝总体重间接决定系数最大的是壳高通过壳宽, 对荣成海域栉孔扇贝则是壳宽通过壳长间接决定总体重。因此, 以总体重为指标挑选青岛海域的种贝时, 可以壳高为优先选择目标, 其次是加强壳宽的协同选择; 挑选荣成海域的种贝时, 可优先选择壳长, 同时加强壳宽的协同选择; 挑选大连海域的种贝时, 可优先选择壳宽, 同时加强壳高的协同选择。
| 类别 | 壳尺寸性状 | 壳长 | 壳宽 | 壳高 |
| 青岛 | 壳长 | 0.036 | 0.080 | 0.111 |
| 壳宽 | — | 0.187 | 0.217 | |
| 壳高 | — | — | 0.199 | |
| 荣成 | 壳长 | 0.358 | 0.237 | 0.139 |
| 壳宽 | — | 0.077 | 0.049 | |
| 壳高 | — | — | 0.020 | |
| 大连 | 壳长 | 0.022 | 0.102 | 0.116 |
| 壳宽 | — | 0.189 | 0.272 | |
| 壳高 | — | — | 0.171 |
各个参数对总体重的回归分析见表 5。对偏回归系数的统计表明, 大连海域栉孔扇贝壳长对总体重的偏回归系数不显著, 为0.266, 其它参数对总体重均能产生极显著的影响(P < 0.01), 说明所有自变量和因变量之间都有极显著的共线性关系。针对青岛、荣成海域的栉孔扇贝可以建立以壳长、壳高和壳宽等壳尺寸性状为自变量, 总体重为因变量的回归方程; 大连海域的栉孔扇贝因其壳长性状影响不显著, 剔除该性状后, 建立以壳高和壳宽为参数的回归方程和回归系数如下:
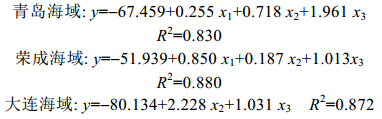
| 模型 | 偏回归系数 | 回归系数 | t | Sig. | ||
| B | 标准误差 | |||||
| 青岛 | (常量) | -67.459 | 3.413 | -19.765 | 0.000 | |
| 壳长 | 0.255 | 0.049 | 0.190 | 5.238 | 0.000 | |
| 壳高 | 0.718 | 0.062 | 0.446 | 11.638 | 0.000 | |
| 壳宽 | 1.961 | 0.150 | 0.432 | 13.086 | 0.000 | |
| 荣成 | (常量) | -51.939 | 2.204 | -23.569 | 0.000 | |
| 壳长 | 0.850 | 0.065 | 0.598 | 13.108 | 0.000 | |
| 壳高 | 0.187 | 0.053 | 0.142 | 3.492 | 0.000 | |
| 壳宽 | 1.013 | 0.122 | 0.277 | 8.301 | 0.000 | |
| 大连 | (常量) | -80.134 | 5.309 | -15.093 | 0.000 | |
| 壳宽 | 2.228 | 0.286 | 0.460 | 7.799 | 0.000 | |
| 壳高 | 1.031 | 0.114 | 0.534 | 9.055 | 0.000 | |
| 注: B为回归方程的常数项, Sig.为差异显著性水平值 | ||||||
多元线性回归的方差分析结果(表 6)指出总体重与壳尺寸性状之间的回归达到极显著水平(P < 0.001)。其中F值分别为383.026、545.539和197.182, 均符合P < 0.01, 方差分析显著, 说明方差分析是有意义的, 可以应用于实际的生产中。
| 海域 | 方差来源 | 平方和 | df | 均方 | F值 | P值 |
| 青岛 | 回归 | 15219.662 | 3.000 | 5073.221 | 383.026 | 0.000 |
| 残差 | 3125.847 | 236.000 | 13.245 | |||
| 总计 | 18345.509 | 239.000 | ||||
| 荣成 | 回归 | 8680.973 | 3.000 | 2893.658 | 545.539 | 0.000 |
| 残差 | 1182.841 | 223.000 | 5.304 | |||
| 总计 | 9863.814 | 226.000 | ||||
| 大连 | 回归 | 14218.440 | 3.000 | 4739.480 | 197.182 | 0.000 |
| 残差 | 2091.136 | 87.000 | 24.036 | |||
| 总计 | 16309.576 | 90.000 | ||||
| 注: df为自由度, P值代表显著性水平 | ||||||
栉孔扇贝属我国北方常见经济贝类, 其一般的养殖周期较短, 为18—24个月, 生长速度存在较明显的季节与年度差异。栉孔扇贝在第一年的生长占一生中生长的29.9%, 第二年的生长占35.2%, 第三年占19.2%, 第四年占8%, 第五年占7.6%, 第二年的生长率在一生中是最大的, 之后逐年递减, 生长缓慢(张玺等, 1956)。本研究选取具有最大生长潜力的二龄到三龄贝开展实验, 以保证其在实验期间能最大限度展现不同养殖环境造成的差异, 同时三龄贝也是种贝生产上的通用规格。值得注意的是, 本研究中仅针对二龄到三龄亲贝的挑选提出指导性建议, 而近年来选育种工作中, 一龄亲贝也可以参与到繁殖群体中, 本结论并不适用于一龄亲贝的挑选工作(作者未发表数据)。
生物的形态性状受遗传和环境共同作用(黄英姿等, 1992), 为尽量减少苗种来源、遗传背景等对本实验的影响, 实验中使用具有相同遗传背景的11(雌)×10(雄)栉孔扇贝混合家系, 并在相同的育苗和养殖环境下养殖一年后, 在扇贝具有相对较大生长潜能(12月龄)时打孔标记, 进行不同纬度海域养殖实验, 从而最大程度降低因初期生长环境和遗传背景不同造成的误差, 使得实验结果说服力更强。
3.2 不同养殖环境下壳尺寸性状对总体重的影响本研究结果表明, 不同养殖环境下具有相同遗传背景的栉孔扇贝混合家系表现出不同的体型特征。不同养殖海域生物的外部形状对总体重的影响不尽相同(毛婷婷等, 2012; 李鸿鹏等, 2015)。于德良等(2013)针对牟平、莱州、广鹿岛、獐子岛四个虾夷扇贝群体的通径分析表明, 均是壳高对活体重量的直接作用最大, 与本文结果有所差异, 可能除了种类不同的原因外, 还和四个群体材料是当地随机选取, 遗传背景并不相同形成差异有关。本实验中影响青岛海域栉孔扇贝总体重的性状按其程度由大到小依次为壳高、壳宽、壳长; 影响荣成海域栉孔扇贝总体重的性状是壳长、壳宽、壳高; 而影响大连海域栉孔扇贝总体重的性状则依次为壳宽、壳高、壳长。本实验中设置不同养殖环境, 主要选取了三个具有一定纬度差别, 又都适合于栉孔扇贝生长的海域。纬度造成的差异主要是温度和日照时间不同, 纬度越高, 温度越低(孙儒泳等, 2002)。生物的生长发育要在达到特定温度时才会开始, 这个温度被称为生物学零度, 生物发育速度随着生物学零度以上的温度增加而增加, 呈线性关系, 并且需要一定时间的积累才可以完成, 这个数值称为有效积温(孙儒泳等, 2002)。不同养殖海域环境下栉孔扇贝生长的有效积温不同。以生物学零度为6°C计, 在一周年的养殖中, 青岛海域栉孔扇贝的有效积温为3075.63°C/d, 荣成海域为2464.34°C/d, 大连海域则为2076.90°C/d(作者未发表数据), 有效积温的不同导致生长和组织积累的差异。由本实验结果可以看出, 亲贝养成海域不同, 在对亲贝进行挑选和分级时其策略亦需要调整。
3.3 影响性状主要因子的确定以及研究的不足之处在使用SPSS进行通径分析时, 相关性指数R2大于或者等于单参数决定系数以及多参数共同决定系数之和时, 才能确定影响因变量的自变量(刘小林等, 2002)。本研究中R2与决定系数之和相等, 表明选用的以上几个参数是影响栉孔扇贝生长的主要关键性状, 通径分析结果能很好的反映壳尺寸性状和总体重之间的关系。并且可用壳尺寸性状对总体重建立多元回归方程, 指导实际生产中亲贝的挑选和分级工作。
值得提出的是, 即使在同一个养殖海域, 环境亦可能存在一定差异, 如较大面积的桑沟湾(133km2), 其不同区域的海流大小、水交换程度明显不同, 营养盐水平和叶绿素a的浓度都有差异(张继红等, 2010; 吴文广等, 2016), 从而初级生产力水平不同。故本研究的结论只适用于以上研究局部区域的浮筏养殖, 并不能一概而论扩大到较大的海域。
4 结论本研究测量了同一来源的栉孔扇贝幼贝在不同纬度海域养殖生长一年后的壳尺寸性状, 并开展了这些壳尺寸形状对总体重性状的通径分析。由于环境的不同, 栉孔扇贝幼贝生长策略存在差异, 故针对不同养殖海域2龄亲贝的挑选工作亦应区别对待。青岛、荣成和大连三个养殖海域栉孔扇贝壳尺寸性状对总体重之间回归方程分别为: y=-67.459+0.255x1+ 0.718x2+1.961x3(R2=0.830), y=-51.939+0.850x1+0.187x2+ 1.013x3(R2=0.880), y=-80.134+2.228x2+1.031x3(R2=0.872)。根据通径分析结果, 对青岛海域栉孔扇贝总体重直接作用最大的是壳高, 对荣成海域扇贝总体重直接作用最大的是壳长, 对大连海域扇贝总体重直接作用最大的是壳宽。因此, 当对三个养殖海域2龄扇贝以总体重为目标进行亲本选择时, 青岛海域应以壳高为优先选择指标; 荣成海域应优先选择壳长; 而大连海域则优先选择壳宽。不同养殖海域栉孔扇贝的通径分析为栉孔扇贝亲贝的挑选提供了理论支持。
| 于瑞海, 王如才, 梁英, 等, 1992. 栉孔扇贝升温育苗高产技术. 海洋湖沼通报, (2) : 68–71 |
| 于瑞海, 包振民, 王昭萍, 等, 2006. 栉孔扇贝×虾夷扇贝的杂交技术. 中国海洋大学学报, 36 (1) : 85–88 |
| 于德良, 丁君, 郝振林, 等, 2013. 不同养殖群体虾夷扇贝数量性状的相关性与通径分析. 大连海洋大学学报, 28 (4) : 350–354 |
| 王冲, 2013. 栉孔扇贝不同性别间重要经济性状比较及通径分析. 水产科学, 32 (8) : 441–446 |
| 王子臣, 1977. 栉孔扇贝的人工育苗与试养. 动物学杂志, 7 (4) : 6–9 |
| 王如才, 高洁, 1986. 栉孔扇贝Chlamys farreri (Jones et Preston)人工育苗试验报告. 山东海洋学院学报, 16 (4) : 145–153 |
| 王如才, 楼伟风, 徐家敏, 等, 1989. 栉孔扇贝氨基酸组份与含量的比较研究. 青岛海洋大学学报, 19 (S2) : 12–19 |
| 王志铮, 吴一挺, 杨磊, 等, 2011. 日本沼虾(Macrobrachium nipponensis)形态性状对体重的影响效应. 海洋与湖沼, 42 (4) : 612–618 |
| 王秀华, 王崇明, 李筠, 等, 2002. 胶州湾栉孔扇贝大规模死亡的流行病学调查. 水产学报, 26 (2) : 149–156 |
| 王玲玲, 方建光, 毛玉泽, 等, 2007. 栉孔扇贝秋季苗种培育、生长特性的初步研究. 海洋水产研究, 28 (5) : 60–65 |
| 毛婷婷, 姜振峰, 李文滨, 2012. 不同遗传背景和环境条件下大豆油分含量与产量性状的相关性和通径分析. 大豆科学, 31 (5) : 744–748 |
| 平洪领, 李玉全, 2013. 逐步线性回归法实现天津厚蟹(Helice tientsinensis)表型性状与体重的通径分析. 海洋与湖沼, 44 (5) : 1353–1357 |
| 刘小林, 常亚青, 相建海, 等, 2002. 栉孔扇贝壳尺寸性状对活体重的影响效果分析. 海洋与湖沼, 33 (6) : 673–678 |
| 孙振兴, 刘相全, 常林瑞, 2013. 基于数量性状的砂海螂(Mya arenaria)选择指标评估. 海洋与湖沼, 44 (3) : 763–767 |
| 孙振兴, 常林瑞, 徐建鹏, 2010. 扁玉螺(Neverita didyma)表型性状对体重和软体部重的影响效应分析. 海洋与湖沼, 41 (4) : 513–518 |
| 孙儒泳, 李庆芬, 牛翠娟, 等, 2002. 基础生态学. 北京: 高等教育出版社, 17-33 |
| 杜美荣, 王彬, 张继红, 等, 2012. 一龄栉孔扇贝壳长与壳高对湿重的相关性和通径分析. 中国农学通报, 28 (20) : 136–139 |
| 杜美荣, 方建光, 毛玉泽, 等, 2009. 栉孔扇贝春季升温育苗与保苗技术研究. 南方水产, 5 (2) : 40–44 |
| 李鸿鹏, 富裕, 任夙艺, 等, 2015. 舟山近海日本囊对虾野生群体与越冬养成群体形态性状对体重和肉重影响的比较. 海洋与湖沼, 46 (5) : 1218–1227 |
| 吴文广, 张继红, 王巍, 等, 2016. 桑沟湾春季叶绿素a浓度分布及其影响因素. 生态学报, 36 (15) : 1–9 |
| 宋微波, 王崇明, 王秀华, 等, 2001. 栉孔扇贝大规模死亡的病原研究新进展. 海洋科学, 25 (12) : 23–26 |
| 张玺, 齐忠彦, 李洁民, 1956. 栉孔扇贝的繁殖和生长. 动物学报, 8 (2) : 235–250 |
| 张永泉, 白庆利, 谷伟, 等, 2014. 白斑红点鲑(Salvelinus leucomaenis)形态性状对体重影响效果. 海洋与湖沼, 45 (3) : 554–560 |
| 张伟杰, 常亚青, 丁君, 等, 2013. 日本镜蛤(Dosinia japonica Reeve)壳尺寸与重量性状的相关与回归分析. 海洋与湖沼, 44 (3) : 796–800 |
| 张根芳, 张文府, 方爱萍, 等, 2014. 养殖条件下不同年龄背瘤丽蚌(Lamprotula leai)数量性状的相关与通径分析. 海洋与湖沼, 45 (5) : 1115–1121 |
| 张继红, 蒋增杰, 王巍, 等, 2010. 桑沟湾营养盐时空分布及营养盐限制分析. 渔业科学进展, 31 (4) : 16–25 |
| 张福绥, 杨红生, 1999a. 栉孔扇贝大规模死亡问题的对策与应急措施. 海洋科学, 23 (2) : 38–42 |
| 张福绥, 杨红生, 1999b. 山东沿岸夏季栉孔扇贝大规模死亡原因分析. 海洋科学, 23 (1) : 44–47 |
| 栗志民, 刘志刚, 王辉, 等, 2011. 企鹅珍珠贝(Pteria penguin)主要经济性状对体重的影响效果分析. 海洋与湖沼, 42 (6) : 798–803 |
| 高玮玮, 袁媛, 潘宝平, 等, 2009. 青蛤(Cyclina sinensis)贝壳形态性状对软体部重的影响分析. 海洋与湖沼, 40 (2) : 166–169 |
| 黄英姿, 毛盛贤, 1992. 基因型与环境互作研究的新进展. 作物学报, 18 (2) : 116–125 |
| 魏利平, 2000. 山东省扇贝养殖现状及持续发展的技术措施. 齐鲁渔业, 17 (2) : 21–23 |
| 魏利平, 翟慎注, 仇宝志, 1985. 栉孔扇贝育苗技术的探讨. 海洋渔业, (1) : 26–29 |
| Guo X M, Ford S E, Zhang F, 1999. Molluscan aquaculture in China. Journal of Shellfish Research, 18 (1) : 19–31 |
| Wright S, 1984. Evolution and the Genetics of Populations Ⅲ. Experimental Results and Evolutionary Deductions. Chicago: University of Chicago Press, 456-461 |