 2017, Vol. 48
2017, Vol. 48中国海洋湖沼学会主办。
文章信息
- 周银环, 曹伏君, 罗杰, 叶宁. 2017.
- ZHOU Yin-Huan, CAO Fu-Jun, LUO Jie, YE Ning. 2017.
- (Boleophthalmus pectinirostris) 耗氧率 (RO) 和排氨率 (RN) 的影响
- INFLUENCE OF TEMPERATURE AND BODY WEIGHT ON OXYGEN CONSUMPTION AND AMMONIA EXCRETION OF BOLEOPHTHALMUS PECTINIROSTRIS
- 海洋与湖沼, 48(2): 392-397
- Oceanologia et Limnologia Sinica, 48(2): 392-397.
- http://dx.doi.org/10.11693/hyhz20161100252
-
文章历史
- 收稿日期:2016-11-18
- 收修改稿日期:2016-12-04




大弹涂鱼 (Boleophthalmus pectinirostris), 又名花跳、跳跳鱼, 隶属于鲈形目 (Perciformes)、弹涂鱼科 (Periophthalmidae) (洪万树等, 1988), 常见于我国东南部沿海的红树林湿地地区, 其肉质鲜美, 营养丰富, 富含多种氨基酸, 具有滋补强身功效, 有较高的经济价值和良好的养殖前景。我国国内对大弹涂鱼的养殖和繁育研究已取得较好成果 (吴善, 2006; 张其永等, 2006), 并获得育苗成功。叶启旺等 (2006)研究了大弹涂鱼土池育苗技术, 其人工养殖正处于推广阶段。大弹涂鱼具有鱼病少、养殖技术容易掌握、活鱼耐长途运输、养殖经济效益高等特点, 促进大弹涂鱼养殖迅猛发展。但目前关于大弹涂鱼其他方面的研究不多, 主要集中在大弹涂鱼的形态特征、行为习性、性腺发育等。刘至治等 (2009)应用RAPD技术及细胞色素b (Cyt b) 基因序列, 分析了长江口及其南部邻近地区大弹涂鱼群体遗传结构及种群历史; 张春丹等 (2006)研究了重金属镉胁迫条件下大弹涂鱼外周血微核率和肝脏过氧化物酶活性的变化。本研究采用室内实验生态学方法, 以大弹涂鱼为研究对象, 对其在不同温度梯度及规格体重下的耗氧率 (RO) 和排氨率 (RN) 变化加以分析, 以期研究温度梯度和规格体重这两项特质对该鱼种的呼吸和排泄的影响作用; 同时也在探究大弹涂鱼能源物质的变化规律, 为对其今后的生理生态、培育繁殖、人工育苗提供理论资料, 促进大弹涂鱼养殖业的可持续发展。
1 材料与方法 1.1 材料实验用鱼是从湛江自然海区当天采捕的大弹涂鱼 (Boleophthalmus pectinirostris)。选择无病无伤、行动迅速、生命力强的个体作为试验用鱼, 按体长分为大、中、小三组 (表 1), 以备实验。pH为 (8.2±0.3), 盐度为 (23.4±0.5), 水温为 (26±0.5) ℃。
| 试验组 | 体长 (cm) | 体重 (g) |
| L | 22.61±2.71 | 21.06±2.86 |
| M | 15.05±0.7 | 12.4±1.00 |
| S | 7.37±0.43 | 9.94±1.06 |
实验设5个温度 (T) 梯度 (16、21、26、31、36) ℃, 每个温度组选三种个体 (大, 中, 小) 进行代谢实验。预实验结果表明, 每组放一尾鱼较为合适, 对每一种个体、各温度梯度均设置2个重复组; 各温度组设置1个无鱼组, 用于溶氧及氨氮测定的对照组。当盐度值为27时, 采用水浴锅和冰块调节温度, 在达到预定的实验温度梯度 (16、21、26、31、36) ℃后, 让鱼在此温度下适应2h。实验代谢瓶 (500mL) 内装100mL海水, 液体石蜡密封。实验持续2h后, 采用虹吸法取代谢瓶中的水样, 测定其溶解氧和氨氨浓度。实验结束后用滤纸将鱼外表的水分吸干, 游标卡尺测量其体长, 电子天平称体重。
1.3 实验计算DO的测定:采用改进的温克勒碘量法 (Winkler法) (GB7489-87); NH4+-N测定:采用奈氏试剂法 (国家环境保护总局, 2002)。并据此计算出耗氧率 (RO) 和排氨率 (RN) 的数值。
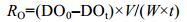
式中, RO为耗氧率[mg/(g.h)]; DO0为空白组流出的海水溶解氧值 (mg/L), DOt为实验大弹涂鱼呼吸流出的水样溶解氧值 (mg/L), V为实验水流速度 (L/h), W为大弹涂鱼体重 (g)。
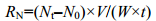
式中, RN为排氨率[μg/(g.h)]; N0为空白组流出的海水氨氮值 (μg/L); Nt为实验大弹涂鱼呼吸后流出的水样氨氮值 (μg/L); V为实验组水流的速度 (L/h); W为大弹涂鱼体重 (g)。
温度系数Q10和氧氮比 (O:N) 的计算方法: Q10的定义为:当水温升高10℃时, 鱼类或其他水产动物的呼吸和排泄发生变化的比率。
公式为: lgQ10 = 10×(lgR2–lgR1)/(T2–T1), 其中R1、R2分别为温度T1、T2的耗氧率或排氨率。
氧氮比 (O:N) 为鱼体消耗氧的克原子数与排出氮克原子数的比率, 即O:N=RO/RN。
1.4 数据的处理对所有数据采用统计软件 (Sta-Tistica 6.0) 进行统计分析。采用Duncan’s多重比较分析组间差异, P < 0.05表示差异显著水平, P < 0.01表示差异极显著水平。
2 结果 2.1 不同温度梯度及规格体重对大弹涂鱼耗氧率 (RO) 的影响如图 1所示, 当温度16—31℃时, 温度对大弹涂鱼的耗氧率有较为显著的影响, 耗氧率随温度的升高而升高, 并在31℃时达到最大值。当水温继续升高时 (31—36℃), 大弹涂鱼的耗氧率下降。在同一温度下, 随着大弹涂鱼体重的增大, 其单位体重的耗氧率逐渐降低。
 |
| 图 1 不同温度梯度和规格体重对大弹涂鱼耗氧率 (RO) 的影响 Fig. 1 Influences of different temperature and weight sizes on RO of B. pectinirostris |
大弹涂鱼的耗氧率 (RO) 与体重 (W) 的回归关系可用幂函数RO=aWb (R2=0.966, P < 0.01) 来表示, 回归方程的系数及相关参数见表 2。表明大弹涂鱼体重与耗氧率呈负相关关系, 随着体重的增加大弹涂鱼的耗氧率呈下降的趋势。
| 温度 (℃) | a | b | R2 | P |
| 16 | 2.546 | –1.207 | 0.947 | < 0.01 |
| 21 | 5.985 | –5.248 | 0.889 | < 0.01 |
| 26 | 2.929 | 0.064 | 0.998 | < 0.01 |
| 31 | 4.146 | –1.131 | 0.996 | < 0.01 |
| 36 | 3.730 | –0.912 | 0.998 | < 0.01 |
方差分析 (ANOVA) 显示 (表 3), 温度和体重对耗氧率有显著影响 (P < 0.01), 而温度和体重的交替作用对耗氧率影响不显著 (P < P0.05)。
| 变异来源 | 自由度 (K) | 平方和 (SSR) | 均方差 (MSR) | 检验量 (F) | 显著性概率 (P) |
| 温度 | 4 | 220692.834 | 55173.208 | 26.683 | < 0.01 |
| 体重 | 2 | 179328.810 | 89664.405 | 43.363 | < 0.01 |
| 温度×体重 | 8 | 25647.335 | 3205.917 | 1.550 | 0.221 |
| 误差 | 15 | 31016.116 | 2067.741 | ||
| 总变异 | 30 | 2130720.955 |
如图 2所示, 大弹涂鱼的排氨率随着温度的升高呈现上升的状态, 低温时, 排氨率呈现缓慢增加的趋势, 在31—36℃时上升的最快。在相同的温度下, 随着大弹涂鱼体重的增加排氨率下降, 而排氨率的变化范围在64.26—496.76µg/(g.h) 之间, 平均为243µg/(g.h)。大弹涂鱼的体重 (W) 与单位体重的排氨率 (RN) 之间的关系符合幂函数方程RN=cWd, 呈负相关关系 (见表 4)。
 |
| 图 2 不同温度梯度和规格体重对大弹涂鱼排氨率 (RN) 的影响 Fig. 2 Influences of different temperature and weight sizes on RN of B. pectinirostris |
| 温度 | c | d | R2 | P |
| 16 | 52.435 | 16.722 | 0.975 | < 0.01 |
| 21 | 83.375 | 25.171 | 0.864 | < 0.01 |
| 26 | 119.73 | 4.8532 | 0.983 | < 0.01 |
| 31 | 113.11 | 65.258 | 0.980 | < 0.01 |
| 36 | 142.44 | 84.583 | 0.967 | < 0.01 |
方差分析 (ANOVA) 显示 (表 5), 温度和体重对排氨率有显著影响 (P < 0.01), 而温度和体重的交替作用对排氨率影响也有显著影响 (P < P0.01)。
| 变异来源 | 自由度K | 平方和SSR | 均方差MSR | 检验量F | 显著性 概率P |
| 温度 | 4 | 39.695 | 9.924 | 29.327 | < 0.01 |
| 体重 | 2 | 307.724 | 153.862 | 454.693 | < 0.01 |
| 温度×体重 | 8 | 40.108 | 5.014 | 14.816 | < 0.01 |
| 误差 | 15 | 5.076 | 0.338 | ||
| 总变异 | 30 | 1490.314 |
大弹涂鱼在16—36℃时, 呼吸Q10范围为0.798—2.440, 平均值为1.495;而排泄Q10范围在1.239—2.997之间, 平均值为1.798, 以上数值基本在正常数据范畴内波动 (表 6)。
| θ(℃) | S | M | L | |||
| 呼吸 | 排泄 | 呼吸 | 排泄 | 呼吸 | 排泄 | |
| 16—21 | 1.200 | 2.294 | 1.791 | 2.997 | 1.346 | 1.937 |
| 21—26 | 1.440 | 1.919 | 1.831 | 1.303 | 2.440 | 1.665 |
| 26—31 | 1.650 | 1.239 | 1.407 | 1.393 | 1.069 | 2.147 |
| 31—36 | 0.913 | 1.579 | 0.798 | 1.663 | 2.237 | 1.570 |
| 平均 | 1.295 | 1.748 | 1.438 | 1.819 | 1.752 | 1.827 |
| T (℃) | S | M | L | |||||||
| O | N | O:N | O | N | O:N | O | N | O:N | ||
| 16 | 6.78 | 169.13 | 0.0400 | 3.19 | 131.3 | 0.0243 | 1.68 | 64.26 | 0.0261 | |
| 21 | 13.92 | 256.19 | 0.0379 | 4.27 | 230.12 | 0.0186 | 1.95 | 89.4453 | 0.0218 | |
| 26 | 8.91 | 354.88 | 0.0251 | 6.86 | 262.63 | 0.0176 | 3.06 | 115.42 | 0.0209 | |
| 31 | 11.45 | 395.35 | 0.0289 | 3.88 | 309.94 | 0.0125 | 3.16 | 169.13 | 0.0187 | |
| 36 | 10.37 | 496.75 | 0.0209 | 6.35 | 399.74 | 0.0159 | 2.91 | 211.87 | 0.0137 | |
| 平均 | 0.0306 | 0.0178 | 0.1012 | |||||||
 |
| 图 3 不同温度下各规格大弹涂鱼耗氧率 (RO) 与排氨率 (RN) 的比值 Fig. 3 Various ratio of RO to RN of B. pectinirostris at different temperature and weight sizes |
鱼类是变温动物, 易受外界环境温度的影响 (朱祥宇等, 2013)。温度是影响鱼类呼吸和排泄等代谢活动最主要的因子 (徐革锋等, 2014)。在一定的适温范围内, 当环境温度由低到高逐渐增加时, 温度越高, 鱼类所需维持生命活动的各组织器官的活性必须增强, 相应的各种酶活性提高, 引起其基础代谢旺盛, 代谢活动不断加强, 活动强度增大, 外在表现即为耗氧率增加 (宋苏祥等, 1997)。对许多鱼类的研究成果均证实了这一点 (闫茂仓等, 2007; 徐钢春等, 2012; 何亚等, 2014; 邓超准等, 2015)。但是, 在适温范围以外, 鱼体的生理机能会发生显著变化, 引起鱼类代谢水平异常, 甚至一些机能性代谢停止活动, 导致鱼体耗氧率迅速下降 (张倩等, 2007)。
在本研究中, (21—31℃) 水温范围内, 随水温的逐步上升, 大弹涂鱼的耗氧率 (RO) 也明显升高, 与对前述鱼类包括鮸鱼、刀鲚、红鳍东方鲀和星洲红鱼等的研究结果基本一致 (闫茂仓等, 2007; 徐钢春等, 2012; 何亚等, 2014; 邓超准等, 2015), 与鱼类RO随温度上升而升高的普遍规律相吻合, 同时, 也说明该温度范围 (21—31℃) 是大弹涂鱼的适合温度。
温度梯度对鱼类的排氨率 (RN) 影响, 主要具体表现在对鱼体组织代谢速率的影响。当水温范围一定时, 随着水温升高, 鱼体的组织代谢速率加快, 所产出的氨、尿素含量增多; 当超过该水温的上限后, 鱼体的组织代谢进入麻痹状态, 甚至可能出现停止状态, 导致鱼体的排氨率也随之降低 (闫茂仓等, 2008; 何亚等, 2014)。本实验的结果表明, 在温度为 (16—36℃) 时, 大弹涂鱼的排氨率随着温度的升高而明显增加, 这与对其他鱼类包括牙鲆 (Paralichthys alivaceus) (张兆琪等, 1997; 张美昭等, 1999)、犬齿牙鲆 (Paralichthys dentatus) (王波等, 2003)、花尾胡椒鲷 (Plectorhynchus cinctus) (王瑁等, 2001; 李加儿等, 2009)、梭鱼 (Liza haematocheila) (线薇薇等, 2002) 等的研究结果较为相似, 也与鱼类的RN随着温度升高而增高的普遍规律相吻合 (闫茂仓等, 2008)。当水温为 (31—36℃) 时, 大弹涂鱼的耗氧率随温度的上升而降低, 但排氨率随水温上升呈继续升高的趋势。
作为温度变化影响鱼类代谢的一项重要指标, Q10值可反映出鱼体内的反应速率与温度之间的关系。鱼类RO与RN随着水温升高而上升, 每当温度上升10℃时, 鱼类的RO可增加约2.3倍 (温度系数Q10= 2.3)(张兆琪等, 1997)。有研究表明, 在温度 (10.8—21.5℃) 范围内, 大西洋牙鲆 (P. dentatus) 的耗氧率Q10值为2.336 (王波等, 2004)。有学者在研究大眼梭鲈 (Stizostedion vitreum) 20—25℃的排氨率时, 对其排氨率Q10进行计算得出其值为2.57 (Forsberg et al, 1992); 对大菱鲆 (Scophthalmus maximus) 排氨率Q10值的研究结果也表明其值为1.37 (Burel et al, 1996); 条石鲷幼鱼 (闫茂仓等, 2008) 呼吸Q10平均为1.274, 排泄Q10平均为1.585。同样, 在相同温度范围内, 南方鲇幼鱼耗氧率与排氨率的Q10值非常接近 (况莉等, 2001), 表明该鱼呼吸与代谢存在相关性, 与本研究结果变化趋势一致。本研究中, 大弹涂鱼在 (16—36℃) 时呼吸Q10范围为0.798—2.440, 平均值为1.495;而排泄Q10值的范围为1.239—2.997, 平均值为1.798, 基本为正常数据的范畴。
3.2 不同体重规格对大弹涂鱼呼吸和排泄的影响鱼类体重也是影响其呼吸和排泄的一个重要因素, 一般研究报道 (Herrmann et al, 2000; 何亚等, 2014) 认为并证实耗氧率和排氨率与体重之间分别呈幂函数关系。本研究中的结果表明, 大弹涂鱼的体重对其呼吸和排泄确有显著影响。随着大弹涂鱼体重的增加, 其耗氧率和排氨率也随之逐步降低, 大弹涂鱼的耗氧率、排氨率与其体重之间分别呈幂函数关系。该结果与其他鱼类的呼吸代谢研究结果一致 (王瑁等, 2001; 线薇薇等, 2002; 王波等, 2003; 李加儿等, 2009)。
一般来说, 鱼类体重对耗氧量的影响表现为:耗氧量随着体重的增加而逐渐升高, 但体重对耗氧率的影响则会因不同鱼种以及生长发育阶段而不同。根据姜祖辉等 (1999)的研究结果, 水生动物用来直接维持其生命的组织和器官 (如肝脏和肾脏等) 的新陈代谢活力高于其他非直接维持生命的组织 (如脂肪和肌肉等), 耗氧率和排氨率的变化可能与鱼类在生长过程中的组织、脏器的比重有关。在鱼类生长过程中, 肝脏和肾脏的比率逐渐降低, 脂肪和肌肉等积累增加, 会导致鱼类的单位重量RO和RN随着个体增大而降低的现象出现。究其原因, 因为直接维持鱼体生命活动的重要组织和器官 (肝脏和肾脏) 在鱼幼小时占整个体重的比例较大, RO和RN也较高, 但该两项数值会随着鱼类个体的增大而呈相对降低的趋势; 另一个可能的原因为, 在不同生长阶段的仔稚鱼, 因其正处于生长发育的过程中, 生长速度越来越快, 代谢活动的旺盛程度远超出成鱼, 导致代谢率随着体质量的增加而不断上升。
3.3 氧氮比 (O:N) 对大弹涂鱼能源物质的分析氧氮比 (O:N) 被众多研究者用来估计生物体代谢过程中的能源物质的化学本质, 氧氮比值的大小反映出蛋白质、脂肪与碳水化合物提供给生物体能量的比率。有学者认为, 如果完全用蛋白质为氧化基质提供能量的话, 其氧氮比值为7—10 (Mayzaud, 1976); 也有学者研究得出结论认为, 如果以蛋白质和脂肪提供能量, 其氧氮比值为24 (Ikeda, 1974); 而Conover等 (1968)经研究后提出, 随着鱼体中脂类和碳水化合物被利用比例逐渐增加, 其氧氮比逐渐升高; 如果完全以脂肪或碳水化合物提供能量的话, 其氧氮比将为无穷大 (Conover et al, 1968)。我国国内研究者对大西洋牙鲆 (P. dentatus) 和
综上所述, 大弹涂鱼体重 (W) 与单位体重的耗氧率 (RO) 之间的关系符合幂函数方程RO= aWb(R2= 0.966, P < 0.01), 呈负指数相关关系。大弹涂鱼的体重 (W) 与单位体重的排氨率 (RN) 之间的关系符合幂函数方程RN= cWd, 呈负相关关系。不同温度梯度和体重规格对大弹涂鱼耗氧率、排氨率均有显著影响 (F > 0.01)。16—31℃时大弹涂鱼主要以脂肪和碳水化合物为能源, 蛋白质其次。
| 王波, 李继强, 曹志海, 等, 2003. 犬齿牙鲆幼鱼呼吸耗氧率的研究. 海洋科学进展, 21(3): 325–330 |
| 王波, 李继强, 曹志海, 等, 2004. 大西洋牙鲆幼鱼标准代谢的初步研究. 海洋科学进展, 22(1): 62–68 |
| 王瑁, 邱书院, 2001. 花尾胡椒鲷幼鱼内、外源性氮和总氮排泄率的研究. 热带海洋学报, 20(3): 87–92 |
| 邓超准, 黄永春, 陈辉辉, 等, 2015. 体质量和温度对星洲红鱼耗氧率、排氨率和窒息点的影响. 淡水渔业, 45(5): 88–93 |
| 叶启旺, 洪万树, 张其永, 等, 2006. 大弹涂鱼土池育苗技术研究. 海洋科学, 30(4): 1–5, 49 |
| 朱祥宇, 高勤峰, 董双林, 2013. 不同温度对草鱼C、N、P营养要素收支的影响. 水产学报, 37(10): 1521–1526 |
| 刘至治, 杨金权, 王正琦, 等, 2009. 长江口及其南部邻近地区大弹涂鱼种群遗传结构及种群历史分析. 动物学研究, 30(1): 1–10 |
| 闫茂仓, 单乐州, 邵鑫斌, 等, 2007. 盐度和pH对鱼幼鱼耗氧率和氨氮排泄率的影响. 台湾海峡, 26(1): 85–91 |
| 闫茂仓, 单乐州, 谢起浪, 等, 2008. 温度、盐度及体重对条石鲷幼鱼耗氧率和排氨率的影响. 海洋科学进展, 26(4): 486–496 |
| 李加儿, 刘士瑞, 区又君, 等, 2009. 花尾胡椒鲷幼鱼的呼吸和排泄代谢. 南方水产, 5(2): 34–39 |
| 吴善, 2006. 大弹涂鱼的人工养殖技术. 福建水产,(3): 48–50 |
| 何亚, 王华, 王伟, 等, 2014. 温度对不同体质量红鳍东方鲀幼鱼耗氧率和排氨率的影响. 大连海洋大学学报, 29(5): 481–485 |
| 况莉, 谢小军, 2001. 温度对饥饿状态下南方鲇幼鱼氨氮排泄的影响. 西南师范大学学报 (自然科学版), 26(1): 45–50 |
| 宋苏祥, 刘洪柏, 孙大江, 等, 1997. 史氏鲟稚鱼的耗氧率和窒息点. 中国水产科学, 4(5): 100–103 |
| 张倩, 袁重桂, 阮成旭, 等, 2007. 褐菖鲉的耗氧率与温度、盐度和体重的关系. 福建水产,(4): 12–16 |
| 张兆琪, 张美昭, 李吉清, 等, 1997. 牙鲆鱼耗氧率、氮排泄率与体重及温度的关系. 青岛海洋大学学报, 27(4): 483–489 |
| 张其永, 洪万树, 陈仕玺, 等, 2006. 潮间带大弹涂鱼产卵习性和自然受精卵孵化研究. 热带海洋学报, 25(5): 68–73 |
| 张春丹, 黄福勇, 李明云, 等, 2006. 镉胁迫条件下大弹涂鱼 (Boleophthalmus pectinirostris) 外周血微核标记及肝脏过氧化物酶标记的变化. 海洋与湖沼, 37(1): 7–13 |
| 张美昭, 张兆琪, 郑春波, 等, 1999. 牙鲆幼鱼能量代谢的初步研究. 中国水产科学, 6(1): 75–78 |
| 国家环境保护总局, 2002. 水和废水监测分析方法. 4版. 北京: 中国环境科学出版社, 279-281 |
| 线薇薇, 朱鑫华, 2002. 梭鱼标准代谢、内源氮排泄与体重和温度的关系. 青岛海洋大学学报, 32(3): 368–374 |
| 姜祖辉, 王俊, 1999. 菲律宾蛤仔氮、磷代谢的初步研究. 青岛大学学报, 12(2): 68–73 |
| 洪万树, 戴庆年, 张其永, 等, 1988. 大弹涂鱼的早期发育研究. 热带海洋,(2): 1–8 |
| 徐革锋, 王裕玉, 韩英, 等, 2014. 不同温度条件下运动和摄食对细鳞鲑幼鱼代谢模式的影响. 应用生态学报, 25(4): 1156–1162 |
| 徐钢春, 聂志娟, 薄其康, 等, 2012. 水温对刀鲚幼鱼耗氧率、窒息点、血糖及肌肝糖元指标的影响. 生态学杂志, 31(12): 3116–3120 |
| Burel C, Person-Le Ruyet J, Gaumet F, et al, 1996. Effects of temperature on growth and metabolism in juvenile turbot. Journal of Fish Biology, 49(4): 678–692 DOI:10.1111/jfb.1996.49.issue-4 |
| Conover R J, Corner E D S, 1968. Respiration and nitrogen excretion by some marine zooplankton in relation to their life cycles. Journal of the Marine Biological Association of the United Kingdom, 48(1): 49–75 DOI:10.1017/S0025315400032410 |
| Forsberg J A, Summerfelt R C, 1992. Effect of temperature on diel ammonia excretion of fingerling walleye. Aquaculture, 102(1-2): 115–126 DOI:10.1016/0044-8486(92)90294-U |
| Herrmann J P, Enders E C, 2000. Effect of body size on the standard metabolism of horse mackerel. Journal of Fish Biology, 57(3): 746–760 DOI:10.1111/jfb.2000.57.issue-3 |
| Ikeda T, 1974. Nutritional ecology of marine zooplankton. Memoirs of the Faculty of Fisheries, Hokkaido University, 22(1): 1–97 |
| Mayzaud P, 1976. Respiration and nitrogen excretion of zooplankton. Ⅳ.The influence of starvation on the metabolism and biochemical composition of some species.Marine Biology, 37(1): 47–58 |