 2018, Vol. 49
2018, Vol. 49中国海洋湖沼学会主办。
文章信息
- 蒋瑞, 刘必林, 刘华雪, 冯春雷, 张健, 倪震宇, 吴旭干, 张虎, 舒畅, 鲁玉渭. 2018.
- JIANG Rui, LIU Bi-Lin, LIU Hua-Xue, FENG Chun-Lei, ZHANG Jian, NI Zhen-Yu, WU Xu-Gan, ZHANG Hu, SHU Chang, LU Yu-Wei. 2018.
- 三种常见经济虾蟹类眼柄微结构分析
- MICROSTRUCTURES OF EYESTALKS OF THREE COMMON COMMERCIAL SHRIMP AND CRAB SPECIES IN CHINA
- 海洋与湖沼, 49(1): 99-105
- Oceanologia et Limnologia Sinica, 49(1): 99-105.
- http://dx.doi.org/10.11693/hyhz20170100014
-
文章历史
- 收稿日期:2017-01-16
- 收修改稿日期:2017-08-18




2. 农业部外海渔业开发重点实验室 广州 510300;
3. 农业部东海与远洋渔业资源开发利用重点实验室 上海 200090;
4. 上海海洋大学水产与生命科学技术学院 上海 201306;
5. 江苏省海洋水产研究所 南通 226007
2. The Key Laboratory of Open-Sea Fishery Development, Ministry of Agriculture, Guangzhou 510300, China;
3. The Key Laboratory of East China Sea & Oceanic Fishery Resources Exploitation and Utilization, Ministry of Agriculture, Shanghai 200090, China;
4. Science and Technology College of Fishery and Life, Shanghai Ocean University, Shanghai 201306, China;
5. Marine Fisheries Research Institute of Jiangsu, Nantong 226007, China
虾、蟹等甲壳类动物的复眼基部有1对可活动的柄, 称为眼柄(许嘉璐等, 1990)。甲壳类动物种类多、数量大, 是渔业资源的重要组成部分, 广泛分布于世界各大洋和南、北极等海域(刘瑞玉, 1988)。年龄是研究水生动物生活史的必要组成部分(Cassidy, 2008), 也是渔业种群动态模型及渔业资源评估中非常重要的参数之一。在之前的文献中, 人们普遍认为不可能直接准确地对甲壳类动物进行年龄鉴定, 因为在甲壳类蜕壳过程中, 记录年龄信息的钙化生长结构全部丢失并被替换(Hartnoll, 2001)。然而, Kilada等(2012)在实验中发现甲壳类的眼柄不会在蜕壳过程中丢失, 在此基础上, Kilada等(2012)首次使用眼柄对美国龙虾(Homarus americanus)、雪蟹(Chionoecetes opilio)、褐雕虾(Sclerocrangon boreas)和北部虾(Pandalus borealis)三种虾蟹类进行年龄鉴定。随后, 越来越多的学者开始使用眼柄对虾蟹类进行鉴龄(Kilada et al, 2015a, b; Leland et al, 2015; Sheridan et al, 2015; Krafft et al, 2016)。然而, 国内相关研究尚未开始, 本文对三疣梭子蟹(Portunus trituberculatus)、中华绒螯蟹(Eriocheir sinensis)和中国对虾(Penaeus orientalis)三种虾蟹类眼柄微结构进行检测与分析, 并找出三者微结构的异同, 从而为今后鉴定甲壳类年龄提供新的可靠的方法。
1 材料与方法 1.1 材料来源中华绒螯蟹样本为2016年11月从上海海洋大学崇明养殖基地采集, 样本个数5个; 三疣梭子蟹样本为2016年10—12月从芦潮港水产市场采购, 样本个数10个; 中国对虾样本为2016年5—9月采集于江苏南通近海, 样本个数10个。各样本的基本生物学信息如表 1所示。
| 种类 | 编号 | 甲长(体长)(mm) | 甲宽(mm) | 体重(g) |
| 三疣 梭子蟹 | S-1 | 79.2 | 153.3 | 197.3 |
| S-2 | 65 | 13.6 | 115.3 | |
| S-3 | 49.4 | 97.2 | 59.9 | |
| S-4 | 50 | 112 | 64.1 | |
| S-5 | 47.2 | 104.1 | 58.7 | |
| S-6 | 48.9 | 104.9 | 56.7 | |
| S-7 | 49.3 | 107.6 | 60.8 | |
| S-8 | 47.8 | 102.9 | 57 | |
| S-9 | 52 | 111.2 | 70.9 | |
| S-10 | 56.1 | 104.4 | 88.1 | |
| 中华 绒螯蟹 | R-1 | 29.2 | 30.6 | 14.1 |
| R-2 | 28.5 | 29.6 | 13.6 | |
| R-3 | 26.7 | 28.9 | 12.3 | |
| R-4 | 25 | 27.1 | 9.8 | |
| R-5 | 27.8 | 29.4 | 12.9 | |
| 中国 对虾 | D-1 | 182 | 20.9 | |
| D-2 | 213 | 30.7 | ||
| D-3 | 152 | 12.3 | ||
| D-4 | 274 | 77.5 | ||
| D-5 | 313 | 117 | ||
| D-6 | 168 | 23.7 | ||
| D-7 | 183 | 21.9 | ||
| D-8 | 205 | 26.3 | ||
| D-9 | 166 | 19.4 | ||
| D-10 | 167 | 18.5 |
样本依据戴爱云等(1977)测量三疣梭子蟹甲长、甲宽和体重, 依据耿绪云等(2007)测量中华绒螯蟹甲长、甲宽和体重, 依据张乃禹(1985)测量对虾体长和体重。蟹类眼柄取下后, 分别用解剖针和煮沸的方法取出眼柄内有机物, 然后置于75%酒精的离心管中保存。虾类眼柄质地较为柔软, 不便于下一步固定, 因此将其沿纵轴线剪开后再去除内部有机物, 然后置于75%酒精的离心管中保存。
三疣梭子蟹和中华绒螯蟹眼柄的眼柄基部剪平后, 竖直放置于模具中, 倒入调配好的冷埋树脂溶液(压克力粉和液体固化剂按体积比1 : 1.5)包埋(图 1)。由于中国对虾眼柄较软, 无法竖直放置, 故将眼柄剪开后, 平放于模具中(图 2)。包埋好的树脂模具放在阴凉通风处24h待其硬化, 硬化后的树脂块依次用120、240、600、1200、2400和2500目的水磨砂纸打磨, 打磨过程中不断在显微镜下检查, 直到打磨到需要的平面, 将经打磨的树脂薄片抛光研磨面(图 1、图 2)。然后将抛光过的截面朝下, 用热熔胶粘在载玻片上, 使用同样的方法对另一面进行打磨, 直至在显微镜下截面的生长纹清晰可见, 再抛光得到眼柄切片(图 1、图 2)。最后用密封袋将其密封保存, 以备观察眼柄微结构之用。
 |
| 图 1 蟹类眼柄切片制作示意图 Fig. 1 Slicing of crab eyestalk |
 |
| 图 2 虾类眼柄切片制作示意图 Fig. 2 Slicing of shrimp eyestalk |
将制作好的切片置于显微镜下, 在连接有电荷耦合(Charge Coupled Device, CDD)传感器的显微镜400倍下对眼柄的生长纹进行拍照。使用Digimizer图像处理软件对照片进行测量, 将钙化层总宽度除以生长纹个数即得到生长纹平均间距, 每个生长纹细纹间距为每两个生长纹细纹之间的间距。
1.4 数据分析方法采用变异系数(Coefficient of variation, CV)分别检验三疣梭子蟹、中华绒螯蟹和中国对虾各样本生长细纹宽度的差异性, 其计算公式如下:
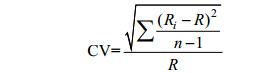
其中, Ri代表第i个样本的间距, R代表样本的平均间距, n表示样本个数。
2 结果 2.1 眼柄前处理方法比较实验发现, 经过煮沸处理之后的眼柄制成的切片在显微镜下观察到的图像不清晰(图 3a), 眼柄内部黑色物质较多, 遮挡住眼柄内部分区域, 影响对眼柄微结构的观察和研究。相比之下, 没有煮沸的眼柄切片观察效果清晰(图 3b)。
 |
| 图 3 煮沸之后的三疣梭子蟹眼柄切片(a)和未煮沸的三疣梭子蟹眼柄切片(b) Fig. 3 The boiled eyestalk slices (a) and unboiled eyestalk slices (b) of P. trituberculatus |
在显微镜下观察三种虾蟹类眼柄结构都分别由4个部分组成(图 4):表层(epicuticle)、色素层(exocuticle)、钙化层(endocuticle)和膜层(membranous layer)。表层是眼柄的最外层, 很薄, 质地比较均匀; 色素层位于表层下方, 是钙化的几丁质层, 内有许多类似黑色素的沉淀物, 在切片磨的很薄的情况下, 可以看到层内有一个个圆柱状物体, 柱状体之间有一定的间隙; 钙化层占整个眼柄厚度的大部分, 是钙化程度最高的一层, 色素较少, 在显微镜的透射光下观察, 有明显的周期性生长纹, 平行于眼柄表层排列, 同时存在垂直于眼柄表层的细纹; 膜层比较薄, 与上两层不同, 不含有钙质, 透明度最高, 是眼柄的生长边缘。
 |
| 图 4 三疣梭子蟹(a)、中华绒螯蟹(b)和中国对虾(c)眼柄切片 Fig. 4 Eyestalk slices of (a) P. trituberculatus, (b) E. sinensis, (c) P. orientalis |
在显微镜下观察三种虾蟹类眼柄都是由4个部分组成, 但是三者眼柄4层微结构特征之间还存在一些差异(表 2), 中华绒螯蟹钙化层含有色素颗粒, 三疣梭子蟹钙化层色素含量很少, 而中国对虾钙化层不含色素。此外, 三种虾蟹类眼柄各层宽度有所不同, 三疣梭子蟹4层分别占总眼柄厚度的比例为6%、17%、64%和13%;中华绒螯蟹4层分别占总眼柄厚度的比例为5%、15%、71%、9%;中国对虾4层分别占总眼柄厚度的比例为6%、16%、57%、21%。
| 种类 | 表层 | 色素层 | 钙化层 | 膜层 |
| 三疣梭子蟹 | 薄 | 较厚, 颜色较深 | 色素含量很少 | 较薄 |
| 中华绒螯蟹 | 薄 | 较厚, 颜色较深 | 含有色素颗粒 | 较薄 |
| 中国对虾 | 薄 | 较薄, 颜色较浅 | 不含色素 | 较厚 |
除钙化层外, 眼柄其他三层结构中鲜有观察到生长纹。三种虾蟹类钙化层里的生长纹大致分为两种(图 5), 第一种是细纹, 纹色浅, 间距小, 排列密, 数量多。越接近钙化层边缘, 细纹之间的间距越小。第二种是宽带, 纹色深, 间距大, 排列疏, 数量少。一个宽带里可以包含着几个或者十几个细纹。三疣梭子蟹眼柄切片中, 钙化区生长纹很清晰, 对比度好, 宽度整体较均匀, 细纹比宽带更清晰, 更容易观察(图 5a)。中华绒螯蟹眼柄钙化层色素较多, 呈颗粒状分布, 不利于观察和读取生长纹(图 5b)。中国对虾钙化层生长纹清晰, 宽带不易观察(图 5c)。
 |
| 图 5 三疣梭子蟹、中华绒螯蟹和中国对虾眼柄钙化层的宽带和细纹 Fig. 5 Bands and increments of endocuticle of (a) P. trituberculatus, (b) E. sinensis, and (c) P. orientalis 注:红点为宽带, 蓝点为细纹 |
实验表明, 三疣梭子蟹眼柄钙化层细纹平均间距为2.9μm, 中华绒螯蟹眼柄钙化层细纹平均间距为4.5μm, 中国对虾眼柄钙化层细纹平均间距为1.2μm。三种虾蟹类眼柄细纹间距从钙化层和色素层的交界到钙化层和膜层的交界都呈现窄-宽-窄的波动变化(图 6), 靠近钙化区边缘部分生长纹间距小, 钙化区中间部分生长纹间距大。中华绒螯蟹眼柄钙化区细纹间距波动最明显, 中国对虾间距波动变化不明显。中华绒螯蟹生长纹最大间距为10.6μm, 三疣梭子蟹生长纹最大间距为6.4μm, 而中国对虾生长纹最大间距只有1.9μm。CV分析结果显示, 三疣梭子蟹、中华绒螯蟹和中国对虾每个个体间眼柄生长纹间距CV值分别为22.2%、26.9%和24.3%。
 |
| 图 6 三疣梭子蟹、中华绒螯蟹和中国对虾生长纹间距 Fig. 6 The space of growth bands of P. trituberculatus, E. sinensis and P. orientalis |
Sheridan等(2015)在观察挪威龙虾(Nephrops norvegicus)眼柄之前, 将其眼柄煮沸, 他认为, 煮沸眼柄能够更容易剔除黏附在眼柄上的有机组织, 使其更好地嵌入冷埋树脂中。但是, 本研究对照经过煮沸和未经煮熟两种眼柄制成的切片后, 认为不将眼柄进行煮沸预处理更有利于观察。虽然煮沸后的蟹类眼柄有机物更易清除, 但煮沸之后的眼柄切片内部黑色物质较多, 遮挡住眼柄内部分区域, 影响对眼柄的观察和研究。而未经煮沸的眼柄切片观察清晰, 虽然存在一些有机组织, 但是在显微镜下观察附着的有机组织仅存在于眼柄膜层外层, 不会遮挡眼柄内部结构, 不影响观察。因此, 在清洗蟹类眼柄之前, 不适宜对其进行煮沸预处理。
甲壳类动物的外壳可分为4个主要部分:表层、色素层、钙化层和膜层(裴思慧等, 2014), 在显微镜透射光下, 三疣梭子蟹、中华绒螯蟹和中国对虾眼柄都分为4个部分, 与赵红霞等(2006)认为的甲壳动物外皮结构是一致的。赵红霞认为表层是由蛋白质和脂类构成, 主要作用与渗透性有关; 色素层里圆柱状物体是充满钙盐的几丁质, 柱状体之间的间隙充实着蛋白质; 膜层则是未钙化的几丁质层。冯玉爱(1984)将甲壳分为2层:非几丁质层和几丁质层, 几丁质层又分为3层——几丁质外层、几丁质中层、几丁质内层, 与本研究观察到的也基本一致。冯玉爱(1984)认为, 在电子显微镜下观察, 几丁质的纤丝状结晶水平排列, 并稀密相间形成明暗分层现象。暗层纤丝密集, 相邻的暗层中有部分纤丝弯向上、下, 稀稀疏疏, 构成明层。部分纤丝弯向上、下排列与本研究中观察到的少量垂直于眼柄表层的细纹一致, 明暗两层也与本研究中观察到的生长纹一致。早在1993年, 林瑛等(1993)在扫描电子显微镜下也发现了甲壳类动物外壳内有平行于壳体表面的层状结构, 主体部分是甲壳质和蛋白质。
冯玉爱(1984)认为虾蟹的甲壳为非细胞结构, 含有丰富的几丁质, 几丁质本身为一种柔软的物质, 但由于其常含有蛋白质和钙质等其他物质, 就变得硬和脆。不同种类的虾蟹、壳的厚薄以及钙质的含量不同, 三疣梭子蟹和中华绒螯蟹眼柄中的钙的含量比中国对虾眼柄中钙含量要多(钙化层厚度比例分别为64%、71%和57%), 因此, 三疣梭子蟹和中华绒螯蟹眼柄硬和较厚, 中国对虾眼柄软而较薄。
一些研究(Kilada et al, 2015a, b; Leland et al, 2015; Sheridan et al, 2015; Krafft et al, 2016)表明, 甲壳类眼柄中的生长纹个数能够反应甲壳类年龄, 认为一个宽带代表 1岁。我们在中国对虾眼柄钙化区中观察到的宽带只有1—2个, 而中国对虾寿命也只有1—2岁(韩光祖等, 1989), 因此, 本研究猜测该宽带个数与中国对虾年龄相关。三疣梭子蟹和中华绒螯蟹眼柄中的宽带多达10个以上, 而两者最大年龄为3—4岁(董志国, 2012; 程泽龙, 2014), 因此本研究认为蟹类眼柄中的宽带个数并不能代表其年龄, 其是否与蜕壳次数有关需要进一步验证。与过去的研究(Kilada et al, 2015a, b; Krafft et al, 2016)有所不同, 我们还在三种虾蟹类眼柄的宽带内部观察到细纹, 每1个细纹代表一天或者数天?其周期性是否与饲养环境有关?仍需进一步研究。
本研究发现, 钙化区边缘部分生长纹间距小, 而钙化区中央部分生长纹间距大, 与一些耳石的日龄宽度变化趋势类似(胡贯宇等, 2015), 与耳石类似, 个体不同生长期生长速率的不同造成生长纹宽度不同。靠近色素层的生长纹代表生长早期, 钙化层中部代表幼体期, 靠近膜层代表成体期, 生长纹宽度的变化趋势与三疣梭子蟹(孙颖民等, 1984)、中华绒螯蟹(陈军伟等, 2016)和中国对虾(张乃禹, 1985)的生长速率变化趋势一致, 也说明不同生长期生长纹越宽表示生长速率越快。
有研究表明(冯玉爱, 1984), 眼柄内生长纹是眼柄内部细胞分泌而成, 与鱼类耳石形成原因类似。耳石上生长轮的宽度往往受摄食条件、水温和光照周期的影响(Neilson et al, 1982), 食物条件通常成为影响耳石生长轮宽度的主要因素(宋昭彬等, 2003)。因此, 本研究推测甲壳类眼柄间距也会受到外部环境影响。中华绒螯蟹眼柄生长纹平均间距最大, 可能是由于样本采集与人工养殖场, 饲养种群的生存条件相对比较稳定, 尤其是食物条件比较稳定。摄食营养好, 生长速度快, 而三疣梭子蟹和中国对虾都是野生样本。中国对虾生长纹平均间距小于三疣梭子蟹生长纹平均间距, 可能是由于中国对虾生长速度慢于三疣梭子蟹。
4 结论长久以来, 虾蟹类的鉴龄工作一直没有获得很好的解决办法。直到2012年, Kilada等(2012)首次发现利用眼柄可以研究甲壳类年龄生长。随后, 甲壳类研究者们开始逐渐尝试利用眼柄研究不同物种的年龄与生长(Kilada et al, 2015a, b; Leland et al, 2015; Sheridan et al, 2015; Krafft et al, 2016)。本研究通过观察三种我国常见的经济虾蟹类眼柄微结构, 了解眼柄微结构形态及其组成, 发现生长纹的宽带细纹之分, 测量每个细纹之间的间距, 可以了解虾蟹类生长情况。本研究认为, 通过观察眼柄微结构, 分析微结构中的生长纹规律, 以此来进一步验证虾蟹类眼柄钙化层的宽带与年龄或脱壳次数和细纹与日龄的关系, 为虾蟹类的年龄鉴定提供新的可靠的方法。并通过对其微化学研究, 来探索虾蟹类生长期间的环境、摄食等生活史信息, 从而为虾蟹类生长和渔业资源研究奠定基础。
| 冯玉爱, 1984. 虾蟹的壳及蜕壳. 福建水产,(2): 46–50 |
| 刘瑞玉, 1988. 海南对虾类. 北京: 农业出版社, |
| 许嘉璐, 彭奕欣, 1990. 中国中学教学百科全书:生物卷. 沈阳: 沈阳出版社, |
| 孙颖民, 宋志乐, 严瑞深, 等, 1984. 三疣梭子蟹生长的初步研究. 生态学报, 4(1): 57–64 |
| 宋昭彬, 常剑波, 曹文宣, 等, 2003. 人工饲养和野生草鱼幼鱼耳石微结构的比较研究. 水生生物学报, 27(1): 7–12 |
| 张乃禹, 1985. 中国对虾生长的数理分析. 海洋科学, 9(4): 1–7 |
| 陈军伟, 马旭洲, 王武, 等, 2016. 不同规格中华绒螯蟹母本子代的生长特性比较. 动物学杂志, 51(5): 895–906 |
| 林瑛, 林瑞洵, 1993. 蟹壳中各组分的组态及其存在关系. 化学通报,(6): 37–40 |
| 赵红霞, 2006. 甲壳动物蜕皮调控研究及蜕壳素的使用. 广东饲料, 15(1): 32–35 |
| 胡贯宇, 陈新军, 刘必林, 等, 2015. 茎柔鱼耳石和角质颚微结构及轮纹判读. 水产学报, 39(3): 361–370 |
| 耿绪云, 王雪惠, 孙金生, 等, 2007. 中华绒螯蟹(Eriocheir sinensis)一龄幼蟹外部形态性状对体重的影响效果分析. 海洋与湖沼, 38(1): 49–54 |
| 董志国, 2012. 中国沿海三疣梭子蟹群体形态、生化与分子遗传多样性研究. 上海: 上海海洋大学博士学位论文 http://cdmd.cnki.com.cn/Article/CDMD-10264-1013000140.htm |
| 韩光祖, 刘永昌, 1989. 秋汛(9-12月)渤海中国对虾生长规律的初步研究. 烟台大学学报(自然科学与工程版),(2): 59–68 |
| 程泽龙, 2014. 中华绒螯蟹新品系的研究. 福州: 福建师范大学硕士学位论文 http://cdmd.cnki.com.cn/Article/CDMD-10394-1015521578.htm |
| 裴思慧, 落继先, 邹恩民, 等, 2014. 摘除眼柄对河南华溪蟹蜕皮激素与几丁质酶含量和β-nagase活性的影响. 水产学报, 38(5): 677–683 |
| 戴爱云, 冯钟琪, 宋玉枝, 等, 1977. 三疣梭子蟹渔业生物学的初步调查. 动物学杂志,(2): 30–33 |
| Cassidy K M, 2008. Use of extractable lipofuscin as an age biomarker to determine age structure of ghost shrimp (Neotrypaea californiensis) populations in west coast estuaries. Corvallis, OR: Master's thesis of Oregon State University http://ir.library.oregonstate.edu/jspui/handle/1957/10312 |
| Hartnoll R G, 2001. Growth in Crustacea——Twenty Years on. In: Paula J P M, Flores A A V, Fransen C H J M eds. Advances in Decapod Crustacean Research. Netherlands: Springer |
| Kilada R, Acu a E, 2015a. Direct age determination by growth band counts of three commercially important crustacean species in Chile. Fisheries Research, 170: 134–143 DOI:10.1016/j.fishres.2015.05.026 |
| Kilada R, Agnalt A L, Arboe N H, et al, 2015b. Feasibility of using growth band counts in age determination of four crustacean species in the northern Atlantic. Journal of Crustacean Biology, 35(4): 499–503 DOI:10.1163/1937240X-00002353 |
| Kilada R, Sainte-Marie B, Rochette R, et al, 2012. Direct determination of age in shrimps, crabs, and lobsters. Canadian Journal of Fisheries and Aquatic Sciences, 69(11): 1728–1733 DOI:10.1139/cjfas-2012-0254 |
| Krafft B A, Kvalsund M, S vik G, et al, 2016. Detection of growth zones in the eyestalk of the Antarctic krill Euphausia superba (Dana, 1852) (Euphausiacea). Journal of Crustacean Biology, 36(3): 167–273 |
| Leland J C, Bucher D J, Coughran J, 2015. Direct age determination of a subtropical freshwater crayfish (redclaw, Cherax quadricarinatus) using ossicular growth marks. PLoS One, 10(8): e0134966 |
| Neilson J D, Geen G H, 1982. Otoliths of chinook salmon (Oncorhynchus tshawytscha):daily growth increments and factors influencing their production. Canadian Journal of Fisheries & Aquatic Sciences, 39(10): 1340–1347 |
| Sheridan M, Officer R, O'Connor I, et al, 2015. Investigating the feasibility of using growth increments for age determination of Norway lobster (Nephrops norvegicus) and brown crab (Cancer pagurus). Journal of Crustacean Biology, 35(4): 189–195 |