 2019, Vol. 50
2019, Vol. 50中国海洋湖沼学会主办。
文章信息
- 何烁, 楼琇林, 史爱琴, 厉冬玲, 王隽, 张华国. 2019.
- HE Shuo, LOU Xiu-Lin, SHI Ai-Qin, LI Dong-Ling, WANG Juan, ZHANG Hua-Guo. 2019.
- 东海海域典型赤潮水体反射光谱模拟研究
- SIMULATION AND STUDY OF REMOTE SENSING REFLECTANCE SPECTRA OF TYPICAL ALGAL BLOOMS IN THE EAST CHINA SEA
- 海洋与湖沼, 50(3): 525-531
- Oceanologia et Limnologia Sinica, 50(3): 525-531.
- http://dx.doi.org/10.11693/hyhz20181100272
文章历史
-
收稿日期:2018-11-12
收修改稿日期:2018-12-17




东海原甲藻是我国东海主要的赤潮优势藻种, 近十几年来每年春季都在浙闽沿岸海域形成大规模赤潮, 并经常与米氏凯伦藻等伴生, 形成有毒有害赤潮(夏平等, 2007; Wang et al, 2009; 郭皓等, 2015)。前人对春季东海赤潮形成机制研究表明, 东海原甲藻个体3—4月份在水体次表层“孕育”, 大量增殖后在5—6月份上升到水体表层形成赤潮(陈翰林等, 2006; 周名江等, 2006)。大量现场调查和观测表明, 东海原甲藻除季节性的垂直迁移现象外, 还有昼夜垂直迁移的习性:凌晨前个体从中下层开始往上迁移, 下午14:30左右聚集到海水表层(王金辉, 2003)。垂直迁移能力为甲藻提供更好的生存条件和更强的环境适应性, 使其相比其他微藻更具竞争优势, 利于赤潮发生(齐雨藻等, 1997; 何涛, 2002; 齐雨藻, 2003)。
赤潮是大量赤潮藻细胞在水体表层聚集, 导致水体变色的海洋现象。海洋水色遥感技术是当前海洋赤潮监测的重要技术手段。赤潮水体反射光谱特征是赤潮遥感的基础, 而其与水体中赤潮藻种类及其浓度、分布状态等因素密切相关。例如, 由于东海原甲藻的吸收光谱在蓝绿波段间存在较大斜率, 其与硅藻间的水体反射光谱差异明显(Tao et al, 2013, 2015)。另外, 东海原甲藻赤潮在从次表层孕育到表层爆发的过程中, 水体中赤潮藻细胞是垂直非均匀分布的(戴鑫烽等, 2014), 其垂直迁移导致赤潮主体水层深度的变化, 加之其在可见光波段的吸收特性, 势必改变水体中光学场的分布, 进而影响水体的反射光谱。分析水体反射光谱对赤潮藻种及其垂向分布变化的响应特性, 有益于赤潮生物量估算、叶绿素浓度反演等遥感算法研究。而探究水体反射光谱对赤潮主体水层深度变化的响应, 有助于建立水体反射光谱与赤潮孕育-爆发过程所处阶段之间的联系, 还可为东海原甲藻垂直迁移等现象的卫星遥感观测提供理论依据。
前人在大洋一类水体组分(叶绿素浓度)垂直非均匀分布对海洋光谱影响方面的研究工作表明, 水表面以下叶绿素浓度最大值的存在增加了蓝光波段的遥感反射率, 减小了绿光波段的遥感反射率(Stramska et al, 2005; Kutser et al, 2008; 席颖等, 2010)。对光学性质复杂的二类水体(例如赤潮孕育期间近岸水体)也已有少量研究, 叶绿素浓度垂直非均匀分布主要影响水体反射光谱的黄绿波段, 而对蓝、红波段几乎没有影响(王云飞等, 2010; 马孟枭等, 2014)。但王云飞在实验中将水体叶绿素浓度最大值设置在10m以深, 不符合赤潮在表层爆发时水体叶绿素浓度的垂直分布特征。另外, 由于纯海水在红光波段的强烈吸收作用(Pope et al, 1997), 较深的水层设置增加了海水对红光的吸收, 这可能是造成红光波段反射率数值变化不明显的原因。基于上述原因, 本文主要开展针对东海原甲藻从次表层(10m以浅)向上垂直迁移过程中水体反射光谱特征及其变化规律研究。
本研究中首先通过模型参数化东海原甲藻在水体中的垂直非均匀分布状态, 设计出各种状态下的水体叶绿素浓度剖面, 近似模拟东海原甲藻的垂直迁移过程。利用Hydrolight 5.3水体光学辐射传输计算软件, 数值计算东海原甲藻水体的反射光谱, 通过分析叶绿素浓度垂直分布模型参数对水体反射光谱的影响, 研究东海原甲藻垂直迁移过程中的水体反射光谱特征及其变化规律。
1 模型与方法对于大洋水体, 在真光层范围内, 水体中叶绿素总量与其表层浓度存在非线性相关性(Gordon et al, 1980), 水体中叶绿素浓度随深度的变化规律通常可以用高斯模型来近似表达(Lewis et al, 1983)。对于我国东海海域, 水体光学特性按剖面形态可分为均匀、线性、跃层及高斯等四种垂向分布类型(徐中伟, 2010)。
东海原甲藻赤潮的多次现场调查和观测结果表明, 甲藻个体在水体中呈垂直非均匀分布, 在次表层存在叶绿素浓度最大值, 可以用用高斯分布模型来表征。本文使用由Platt改进后的高斯模型来表征东海原甲藻水体中叶绿素浓度的垂直分布规律(Platt et al, 1988)。水体叶绿素浓度C垂直分布Platt高斯模型数学表达式为:
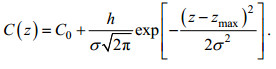 (1)
(1)公式(1)中, C0为背景叶绿素浓度, 单位为mg/m3。zmax为叶绿素浓度峰值所在深度, 单位为m。h为背景叶绿素浓度以上的水体总生物量, 单位为mg/m2。σ为标准方差, 控制着由高叶绿素浓度表征的甲藻主体水层的厚度, 单位为m, 95%的总生物量出现在叶绿素浓度峰值深度±2σ范围内。此外, 水体中叶绿素浓度峰值由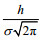
 |
| 图 1 水体叶绿素浓度垂直分布高斯模型 Fig. 1 The Gauss vertical distribution model of chlorophyll concentration 注: C表示叶绿素浓度; C0表示背景叶绿素浓度; h表示水体生物量; zmax表示叶绿素浓度峰值深度; σ表示标准方差 |
为了模拟东海原甲藻在次表层水体中的各种分布状态及其垂直迁移过程, 我们设置zmax位于表层(0m)和次表层(10m)之间, 垂向变动步长为1m。C0共设置四个数值, 分别为0.1、0.5、1.0、1.5mg/m3。σ设置4个数值, 分别为0.8、1.2、1.6、2.0。h设置4个数值, 分别为50、100、150、200mg/m2。这些模拟参数的设置参考了前人研究成果及研究海域现场航次调查结果(王云飞等, 2010; He et al, 2018)。在利用Hydrolight软件进行典型赤潮甲藻水体光谱模拟时, 还需要确定各水体组分(叶绿素、悬浮物和黄色物质)的光学特性参数。考虑到我国东海近岸海域水体的光学复杂性, 本文选用适合浙闽沿岸海域的二类水体生物光学模型。模型中包括对各水体组分吸收、散射及后向散射系数的设置, 其中纯海水的吸收和散射, 选取Pope, Fry和Morel的测量结果(Pope et al, 1997; Morel, 1974); 浮游植物的散射和后向散射概率选取Gordon和Twardowski的模型(Gordon et al, 1980; Twardowski et al, 2007); 黄色物质的吸收选取Bricaud的经典模型; 非藻类颗粒的吸收散射, 后向散射模型分别采用丘仲锋、宋庆君和刘炜的相应研究成果(宋庆君等, 2006; 丘仲峰等, 2007; 刘炜, 2007)。此外, 针对模拟对象东海原甲藻, 模型中藻类吸收系数采用东海原甲藻的吸收光谱测量结果(Tao et al, 2013)。在模拟过程中, 设置水体中非藻类颗粒物和黄色物质垂直均匀分布, 根据前人对东海海域水体组分光学参数的测量结果, 将非藻类颗粒物的背景浓度为1g/m3, 黄色物质在440nm波长处的吸收系数为0.3m–1(宋庆君等, 2006; 朱建华等, 2012; 周虹丽等, 2015)。
为定量描述各实验组中反射光谱的变化程度, 本文采用遥感反射率归一化差值(ΔR)来衡量当模型参数在数值上增加设置的单位步长时, 对应的水体遥感反射率数值的变化幅度(王云飞, 2009):
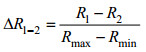
其中Rn(n=1, 2)表示参与比较的该组第n条遥感反射率光谱, 如R1-R2表示该组第1条光谱与第2条光谱对应各波段的数值之差, Rmax表示该组参与比较的两条光谱在所有相同波段遥感反射率变化的正最大值, Rmin为遥感反射率变化的负最大值。
2 结果与分析 2.1 背景叶绿素浓度的影响为分析背景叶绿素浓度对水体反射光谱的影响, 选取水体生物量为200mg/m2, 标准方差为2m, 叶绿素浓度峰值深度为4m, 背景叶绿素浓度为0.1、0.5、1.0、1.5mg/m3的4条光谱曲线如图 2所示。
 |
| 图 2 背景叶绿素浓度C0对水体遥感反射率R的影响 Fig. 2 The effect of chlorophyll background concentration C0 on R 注: R表示遥感反射率, ΔR表示遥感反射率归一化差值 |
由图 2a可见, 在波长400—560nm范围内, 光谱曲线间差异很小, 背景叶绿素浓度低的光谱曲线数值稍大。在560—660nm波长范围内, 光谱间数值差异微小, 不同光谱曲线几乎重合。在660—720nm波长范围内, 光谱间差异也很小, 背景叶绿素浓度低的光谱曲线数值较小。在图 2b中, 波长400—540nm区间, ΔR曲线数值为正, 而在大于540nm的波长区间, ΔR数值几乎全为负值, 前后两个波段区间变化趋势相反。
上述对比分析表明, 较低范围内背景叶绿素浓度的变化对赤潮水体反射光谱的影响不明显, 仅在叶绿素吸收(450 —550nm)及荧光效应波段(675nm)有微弱影响。背景叶绿素浓度在较低范围内变大时, 水体反射光谱数值在蓝绿波段减小, 而在黄红波段增加, 但光谱数值改变不大。
2.2 甲藻主体水层厚度的影响叶绿素浓度高斯分布模型中的标准方差(σ)决定水体中高叶绿素浓度的甲藻主体水层厚度。为探究甲藻主体水层厚度对水体反射光谱的影响, 选择背景叶绿素浓度为1mg/m3, 生物量为200mg/m2, 叶绿素浓度峰值深度为4m, 标准方差为0.8, 1.2, 1.6, 2.0m的4条光谱曲线如图 3所示。
 |
| 图 3 东海原甲藻主体水层深度σ对水体遥感反射率R的影响 Fig. 3 The effect of the depth of Prorocentrum donghaiense Lu layer σ on R |
在图 3a中, 波长430—520nm范围的水体反射光谱曲线数值较为接近, 变化相对较小。在520—720nm波长区间, 光谱曲线之间的数值差异较为明显, 甲藻主体水层厚度越薄的光谱曲线数值也越小, 表明甲藻在水体次表层聚集过程中, 水体反射光谱将变小。在图 3b中可见, 归一化差值ΔR在短波波段数值变化较小, 最大变化出现在ΔR1.2—1.6的686nm附近, 表明甲藻主体水层厚度变化对叶绿素荧光效应波段的遥感反射率影响显著。
以高叶绿素浓度为代表的甲藻主体水层厚度变化对水体反射光谱影响明显, 尤其在荧光效应波段影响显著。在叶绿素浓度峰值深度固定时, 标准方差数值减小, 意味着甲藻主体水层厚度变薄, 甲藻在水体中更集中, 水体叶绿素浓度更高, 光束越容易被吸收, 导致水体反射光谱数值整体变小。对应在荧光效应波段, 光束经过水层内叶绿素强烈的吸收作用后, 散射出海面的荧光光束传输距离相对变大, 被水体吸收更为严重, 使得东海原甲藻水体的荧光效应减弱。
2.3 水体总生物量的影响水体总生物量越大意味着各水层叶绿素浓度越高。为研究水体总生物量对反射光谱的影响, 选择叶绿素浓度峰值深度为4m, 背景叶绿素浓度为1mg/m3, 标准方差为2m, 水体总生物量分别为50、100、150、200mg/m2的水体反射光谱曲线如图 4所示。
 |
| 图 4 水体总生物量h对水体遥感反射率R的影响 Fig. 4 The effect of the total biomass h on R |
由图 4a可见, 在波长400—450nm、580—600nm区间内, 水体反射光谱曲线间数值差异较小, 曲线几乎重合。在450—580nm波长范围, 水体总生物量越大, 水体反射光谱曲线数值越小, 光谱曲线间数值差异明显。在600—720nm波长区间, 总生物量越大, 光谱曲线数值越大。图 4b中, 在400—450nm波长区间, 归一化差值ΔR的数值变化也不明显; 在450—530nm范围, ΔR为正值, 最大变化值出现在480—520nm处; 而在530nm以上波长区间, ΔR均为负值, 其中最大变化出现在总生物量为50—100mg/m2变化曲线的675nm波长处。
水体总生物量改变主要影响水体反射光谱的蓝绿波段和荧光效应波段, 并在560nm波长处出现反转现象。当总生物量增加时, 水体中叶绿素浓度变大, 更多入射光束被水体吸收, 表现为叶绿素主要吸收波段的遥感反射率减小, 同时在长波波段散射出更多的叶绿素荧光, 对应遥感反射率变大。
2.4 叶绿素浓度峰值深度的影响叶绿素浓度峰值深度是甲藻主体水层所在的深度, 它的上下变动是东海原甲藻垂直迁移的最主要表现之一。为了探究叶绿素浓度峰值深度变化对水体遥感反射率的影响, 我们选择背景叶绿素浓度为1mg/m3, 总生物量为200mg/m2, 标准方差设为2m, 叶绿素浓度峰值深度在4—7m范围内变化的水体光谱曲线如图 5所示。
 |
| 图 5 叶绿素浓度峰值深度zmax对水体遥感反射率R的影响 Fig. 5 The effect of depth of chlorophyll maximum zmax on R |
在图 5a中, 在430—450nm、550nm的波长区间, 水体反射光谱间的数值变化不大。在波长450— 550nm范围内, 叶绿素浓度峰值深度越小, 对应光谱曲线的R数值也越小, 相反在550—720nm波长区间, 光谱曲线对应的R数值越大, 这两个波长区间的的光谱曲线差异明显, 变化趋势相反。在图 4b中, 400— 550nm, 归一化差值ΔR大部分为负值, 最大变化值出现在500nm附近。在570—720nm波段区间, ΔR基本为正值, 其中最大变化值出现在ΔR4-5的680nm波长处。
叶绿素浓度峰值深度改变影响水体反射光谱的蓝绿波段和黄红波段, 且变化趋势在550nm波长处反转。叶绿素浓度峰值深度减小, 意味着甲藻主体水层上升, 东海原甲藻从下向上垂直迁移, 表层水体中的叶绿素浓度升高, 水体对光束的吸收作用增强, 使反射光谱曲线蓝绿波段的R数值降低。而叶绿素荧光在向海面传输过程中经过的水层深度变浅, 水体组分对荧光光束的吸收减少, 使得680nm波长处荧光效应波段的反射率数值增加明显。
2.5 东海原甲藻赤潮发生过程模拟分析在2.4节对东海原甲藻垂直迁移现象的模拟研究中, 假设一天内单位水柱内总生物量(包含背景值以上总生物量h与背景叶绿素浓度C0)不发生变化, 则当叶绿素浓度峰值深度zmax小于等于2σ时, 只改变峰值深度zmax的情况下, 可保持单位水柱内总生物量的稳定。但是, 当峰值深度zmax小于2σ时, 对高斯模型而言, 需要改变模型结构才能实现单位水柱内总生物量不变。这种情况发生在东海原甲藻从次表层上升到表层, 甲藻个体在水体表层聚集, 赤潮爆发。东海原甲藻赤潮发生过程中的水体反射光谱模拟结果如图 6所示。
 |
| 图 6 东海原甲藻赤潮发生过程中水体遥感反射率R变化 Fig. 6 The course of R change of the Prorocentrum donghaiense Lu bloom |
从图 6a可见, 当预设C0为1.0mg/m3, σ为2m, h为100mg/m3时, zmax从4m开始依次减小, 甲藻主体水层逐渐上升, 东海原甲藻垂直向上迁移。在只改变zmax的情况下, 当zmax=4时, 水柱内总生物量h4接近预设总生物量h的97.7%, 可认为此时生物量没有变化; 当zmax=3时, 水柱内总生物量h3约为预设值的93%;当zmax=2时, 水柱内生物量h2为预设值的84%, 这时的生物量减少已经较为明显; 当zmax=1时, 水柱内的生物量h1只有预设值的69%, 对应的水体反射光谱如图 6a中No.6、No.5、No.4、No.3所示。随着zmax的减小, 水体反射光谱曲线变化规律与前文基本一致, 其中当zmax=2(No.4)和zmax=1(No.3)时, 两条光谱在短波处的数值较为接近。
考虑到上述接近可能是由于单位水柱内总生物量的减少而造成的, 但这违背了水体总生物量不变的前提。因此, 需要同时改变叶绿素浓度高斯分布模型的其他参数, 来维持水体生物量平衡。以zmax=1为例分析, 要保证水柱内生物量与zmax=2时的一致, 有两种方案:一是增加预设生物量h, 使水柱内的生物量h1同步增大, 从而保证与h2数值相等, 计算表明这种方法需要将预设生物量增加到122mg/m3。二是减小标准方差σ, 当σ减小为1m时, 实际生物量h1与h2一致。此外, 同时改变这两种参数也可以达到目的, 但参数设置过于复杂且不具有代表性, 本文不作讨论。
通过改变标准方差和生物量, 我们得到图 6b中的水体反射光谱曲线No.1和No.2。与曲线No.3比较, 两种方式都增加了表层水体内的叶绿素浓度, 使得光谱曲线在蓝绿波段和荧光波段的数值变化更加明显。综上, 可以看出当甲藻主体水层深度上升为1m时, 叶绿素浓度垂直分布剖面结构发生了明显变化, 东海原甲藻个体在表层聚集, 赤潮爆发。赤潮爆发状态下的水体反射光谱与东海原甲藻向上垂直迁移过程中的光谱特征和变化规律基本一致, 水体反射光谱在蓝绿波段(400—570nm)减小, 在黄红波段(570—720nm)增大。
3 结论本文以东海原甲藻为例, 通过参数化典型赤潮甲藻在水体中的垂向分布状态, 来模拟典型赤潮甲藻垂直迁移过程中的水体叶绿素浓度垂直分布。利用Hydrolight软件数值计算了不同状态下的东海原甲藻水体的反射光谱, 分析了水体反射光谱的影响因素, 研究了水体反射光谱特征, 并探讨了典型赤潮甲藻从次表层向表层迁移到赤潮爆发过程的水体反射光谱变化规律, 得到以下结论:
(1) 叶绿素浓度垂直分布高斯模型中, 模型参数(背景叶绿素浓度、水体总生物量、甲藻主体水层厚度和叶绿素浓度峰值深度)改变对典型甲藻赤潮水体反射光谱有不同程度的影响, 其中背景叶绿素浓度的影响相对较小。
(2) 甲藻主体水层厚度主要影响赤潮水体反射光谱绿黄红波段(520—720nm), 主体水层厚度越薄, 甲藻个体在水体次表层越集中, 水体反射光谱数值越小。
(3) 水体总生物量改变主要影响赤潮水体反射光谱的蓝绿波段和荧光效应波段, 并在560nm波长处出现反转现象。当总生物量增加时, 蓝绿波段的遥感反射率减小, 荧光效应波段增大。
(4) 在东海原甲藻从次表层向表层迁移到赤潮爆发过程中, 赤潮水体反射光谱在蓝绿波段(400—570nm)减小, 在黄红波段(570—720nm)增大, 特别是在叶绿素荧光效应波段(680nm)处的变化尤为明显。
马孟枭, 张玉超, 钱新, 等. 2014. 藻类垂向分布对内陆湖泊叶绿素a反演算法影响的模拟研究. 红外与毫米波学报, 33(6): 666-673 |
王云飞, 2009.东海赤潮监测卫星遥感方法研究.青岛: 中国海洋大学博士学位论文, 88-89
|
王云飞, 贺明霞. 2010. 东海赤潮多发区非均匀叶绿素浓度剖面对遥感反射比的影响. 中国海洋大学学报, 40(10): 100-104 |
王金辉. 2003. 东海赤潮生物具齿原甲藻及其特征. 浙江海洋学院学报(自然科学版), 22(2): 128-131 DOI:10.3969/j.issn.1008-830X.2003.02.008 |
丘仲锋, 唐军武, 何宜军. 2007. 东海赤潮高发区水体光谱吸收特性分析. 海洋科学集刊, 48: 6-19 |
朱建华, 周虹丽, 李铜基, 等. 2012. 中国近海黄色物质吸收光谱特征分析. 光学技术, 38(3): 295-299 |
刘炜, 李铜基, 朱建华, 等. 2007. 黄东海海区总悬浮物散射特性研究. 海洋技术学报, 26(2): 42-46 |
齐雨藻. 2003. 中国沿海赤潮. 北京: 科学出版社
|
齐雨藻, 黄长江, 钟彦, 等. 1997. 甲藻塔玛亚历山大藻昼夜垂直迁移特性的研究. 海洋与湖沼, 28(5): 458-467 |
何涛, 2002.厦门筼筜湖赤潮发生的生态学研究.厦门: 厦门大学硕士学位论文, 29-30
|
宋庆君, 唐军武. 2006. 黄海、东海海区水体散射特性研究. 海洋学报, 28(4): 56-63 |
陈翰林, 吕颂辉, 张传松, 等. 2006. 2004年东海原甲藻赤潮爆发的现场调查和分析. 生态科学, 25(3): 226-230 DOI:10.3969/j.issn.1008-8873.2006.03.009 |
周名江, 朱明远. 2006. "我国近海有害赤潮发生的生态学、海洋学机制及预测防治"研究进展. 地球科学进展, 21(7): 673-679 DOI:10.3321/j.issn:1001-8166.2006.07.003 |
周虹丽, 朱建华, 李铜基. 2015. 中国近海黄色物质吸收光谱经验斜率特征研究. 热带海洋学报, 34(1): 23-29 DOI:10.3969/j.issn.1009-5470.2015.01.004 |
夏平, 陆斗定, 朱德弟, 等. 2007. 浙江近岸海域赤潮发生的趋势与特点. 海洋学研究, 25(7): 47-56 |
徐中伟, 2010.垂直非均匀海洋的海洋光学特性.青岛: 中国海洋大学硕士学位论文, 45-46
|
郭皓, 丁德文, 林凤翱, 等. 2015. 近20a我国近海赤潮特点与发生规律. 海洋科学进展, 33(4): 547-558 DOI:10.3969/j.issn.1671-6647.2015.04.013 |
席颖, 杜克平, 张丽华, 等. 2010. 叶绿素浓度垂直不均一分布对于分层水体表观光学特性的影响. 光谱学与光谱分析, 30(2): 489-494 DOI:10.3964/j.issn.1000-0593(2010)02-0489-06 |
戴鑫烽, 陆斗定, 夏平, 等. 2014. 2010-2011年东海藻华高发区水体层化对东海原甲藻(Prorocentrum donghaiense Lu)藻华的影响. 海洋与湖沼, 45(2): 217-224 |
Gordon H R, Clark D K, 1980. Remote sensing optical properties of a stratified ocean: an improved interpretation. Applied Optics, 19(20): 3428-3430 DOI:10.1364/AO.19.003428 |
He S, Lou X, et al, 2017. Remote sensing reflectance simulation of coastal optical complex water in the East China Sea. Proc. SPIE 10607, MIPPR 2017: Multispectral Image Acquisition, Processing, and Analysis, 106070U.
|
Kutser T, Metsamaa L, Dekker A G, 2008. Influence of the vertical distribution of cyanobacteria in the water column on the remote sensing signal. Estuarine, Coastal and Shelf Science, 78(4): 649-654 DOI:10.1016/j.ecss.2008.02.024 |
Lewis M R, Cullen J J, Platt T, 1983. Phytoplankton and thermal structure in the upper ocean: consequences of nonuniformity in chlorophyll profile. Journal of Geophysical Research: Oceans, 88(C4): 2565-2570 DOI:10.1029/JC088iC04p02565 |
Morel A, Smith R C, 1974. Relation between total quanta and total energy for aquatic photosynthesis. Limnology & Oceanography, 19(4): 591-600 |
Platt T, Sathyendranath S, 1988. Oceanic primary production: estimation by remote sensing at local and regional scales. Science, 241(4873): 1613-1619 DOI:10.1126/science.241.4873.1613 |
Pope R M, Fry E S, 1997. Absorption spectrum (380-700 nm) of pure water. Ⅱ. Integrating cavity measurements. Applied Optics, 36(33): 8710-8723 DOI:10.1364/AO.36.008710 |
Stramska M, Stramski D, 2005. Effects of a nonuniform vertical profile of chlorophyll concentration on remote-sensing reflectance of the ocean. Applied Optics, 44(9): 1735-1747 DOI:10.1364/AO.44.001735 |
Tao B Y, Pan D L, Mao Z H et al, 2013. Optical detection of Prorocentrum donghaiense blooms based on multispectral reflectance. Acta Oceanologica Sinica, 32(10): 48-56 DOI:10.1007/s13131-013-0365-6 |
Tao B Y, Mao Z H, Lei H et al, 2015. A novel method for discriminating Prorocentrum donghaiense from diatom blooms in the East China Sea using MODIS measurements. Remote Sensing of Environment, 158: 267-280 DOI:10.1016/j.rse.2014.11.004 |
Twardowski M S, Claustre H, Freeman S A et al, 2007. Optical backscattering properties of the "clearest" natural waters, Biogeosciences, 4(6): 1041-1058
|
Wang J H, Wu J Y, 2009. Occurrence and potential risks of harmful algal blooms in the East China Sea. Science of the Total Environment, 407(13): 4012-4021 DOI:10.1016/j.scitotenv.2009.02.040 |