 2020, Vol. 51
2020, Vol. 51中国海洋湖沼学会主办。
文章信息
- 刘金虎, 李毅, 曹亮, 许莉莉, 窦硕增. 2020.
- LIU Jin-Hu, LI Yi, CAO Liang, XU Li-Li, DOU Shuo-Zeng. 2020.
- 闽东近海游泳动物群落组成、资源量与生物多样性的时空格局研究
- THE TEMPO-SPATIAL CHANGES IN COMMUNITY STRUCTURE, BIOMASS, AND BIODIVERSITY OF NEKTON IN THE EASTERN COASTAL WATERS OF FUJIAN PROVINCE
- 海洋与湖沼, 51(4): 931-942
- Oceanologia et Limnologia Sinica, 51(4): 931-942.
- http://dx.doi.org/10.11693/hyhz20200100017
文章历史
-
收稿日期:2020-01-14
收修改稿日期:2020-03-17




2. 华能霞浦核电有限公司 宁德 352100;
3. 中国科学院大学 北京 100049
2. Huaneng-Xiapu Nuclear Power Co. Ltd., Ningde 352100, China;
3. The University of Chinese Academy of Sciences, Beijing 100049, China
福建沿海有闽江、九龙江等众多河流输入, 与台湾暖流等交汇于此, 为渔业生物提供了丰富的饵料基础; 众多的河口和海湾曾为多种渔业生物提供了栖息地, 形成了重要优良渔场(戴泉水等, 2005;黄良敏, 2011)。目前, 闽东等近海渔场面临捕捞压力大和生境退化等多压力, 近海渔业生产力下降, 鱼类生活史、种群数量、洄游分布和生物群落结构等发生显著变化, 主要表现为游泳动物种类剧减, 物种多样性降低, 群落和优势种更替明显, 种群小型化问题显著(戴泉水等, 2005;刘勇等, 2006;沈长春, 2011)。20世纪80年代, 闽东近海记录的游泳动物超过300种, 优势种包括大黄鱼、带鱼、银鲳等众多优质经济鱼类。但是, 近年来的调查中出现的游泳动物为100种左右, 优势种为龙头鱼、
近年来, 一些学者开展了福建沿海游泳动物或渔业生物群落特征及资源现状的相关研究。调查海域包括七星列岛、三沙湾、闽江口、东山湾、台湾海峡和九龙江口等海域(戴泉水等, 2005;黄良敏, 2011;张会军, 2013;张静等, 2013;顾洪静, 2014;张春草, 2017;叶孙忠等, 2018)。研究内容涉及渔业生物种类组成、数量分布、群落结构、生物多样性、资源密度空间分布、资源承载力评估等。相关研究多局限于特定的小尺度渔业水域。本研究以沿岸有多个在建或已运营的核电厂的我国传统渔场——闽东渔场为调查水域, 研究游泳动物群落结构特征、资源量和生物多样性时空分布格局, 以期为评价该水域环境变化下渔业资源变动特征和资源养护管理决策提供科学参考。
1 材料与方法 1.1 调查站位与区域于2017年春季(5月)和秋季(10月)在闽东近海开展定点渔业生物资源调查。调查范围位于闽东及台湾浅滩北部的部分区域, 范围为119°44′—121°21′E, 25°43′—27°19′N。调查共设计断面8条, 底拖网站位29个(图 1)。
 |
| 图 1 闽东近海游泳动物调查海区及站位 Fig. 1 Sampling sites of the fishery survey in the eastern coastal waters of Fujian Province |
利用单囊底拖网(渔船功率220kW, 网口长32m, 网长25m, 囊网网目20mm)开展海上调查取样。依据《海洋渔业资源调查规范》(SC/T9403- 2012)相关的要求, 在每个调查站位拖网1h, 拖速2.0kn。同时调查各站位的现场表层水温、盐度、水深等环境因子。拖网结束后, 现场计测渔获量并取样; 在室内对样本进行种类鉴定、计数和生物学测定。
1.2 数据统计分析利用相对重要性指数(Index of relative importance, IRI)判定优势物种(Pianka, 1971):
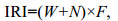 (1)
(1)式中, W为物种在各站位的渔获中的重量百分比, N为个体数量百分比, F为出现频率。IRI > 500为优势种, 100≤IRI < 500为重要种, 10≤IRI < 100为常见种。
利用Shannon-wiener指数(H′)和Pielou均匀度指数(J′)评价各站位的生物多样性(个体数生物多样性和重量生物多样性):
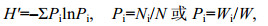 (2)
(2)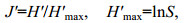 (3)
(3)式(2)和式(3)中, H′为物种多样性指数, H′max为最大物种多样性指数, S为总物种数, Ni或Wi为第i物种的生物个体数量或重量, N或W为所有物种的个体数量或重量总和, Pi为第i物种占所有物种的个体数量或重量百分比。
单位捕捞努力量渔获量(catch per unit effort, CPUE)为每小时单位网次的总渔获量[ind或kg/(net·h)], 据此计算生物资源密度D (ind/km2或kg/km2):
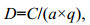 (4)
(4)式(4)中, C为每小时渔获量(ind或kg/h), a为每小时扫海面积(km2), q为可捕系数。头足类、甲壳类和鱼类的可捕系数分别取0.5、0.8和0.7(林龙山等, 2012)。
2 结果 2.1 种类组成春、秋季调查中, 共出现游泳动物70种, 隶属于15目40科64属(见附录)。其中, 头足类3目4科4属5种, 占总捕获物种数的7.1%;甲壳类2目9科15属18种, 占25.7%;鱼类10目27科45属47种, 占67.1%。大多数为定居性种。
甲壳类包括十足目种类18种和口足目1种(口虾蛄)。十足目中出现对虾科4种和梭子蟹科3种, 其他科种类数较少。软骨鱼纲中仅出现孔鳐1种, 45种硬骨鱼类主要有鲈形目鱼类(25种), 以石首鱼科(6种)和
春季渔获中出现56种, 隶属于13目32科51属, 包括头足类4种、甲壳动物16种和鱼类36种。秋季出现59种, 隶属15目35科56属, 包括头足类5种、甲壳动物14种和鱼类40种。
2.2 优势种组成春季, 各物种IRI值范围为1—5263, 优势种有红狼牙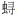
在春、秋两季中, 三疣梭子蟹、日本
春季, 鱼类、甲壳类和头足类个体数量分别占总渔获个体数量的66.4%、33.4%和0.2%, 其个体重量分别占总渔获重量的72.3%、26.9%和0.8%。秋季, 三个生物类群在总渔获个体数量中的占比分别为56.6%、42.0%和1.4%, 个体重量占比分别为60.4%、37.8%和1.8% (表 1)。
| 优势种/生物类群 | 春季 | 秋季 | ||
| 个体数量比N(%) | 个体重量比W(%) | 个体数量比N(%) | 个体重量比W(%) | |
| 三疣梭子蟹 | 9.4 | 14.0 | 18.2 | 25.5 |
| 日本蟳 | 4.6 | 1.7 | 5.0 | 3.5 |
| 龙头鱼 | 3.6 | 8.9 | 22.1 | 24.1 |
| 棘头梅童鱼 | 3.3 | 4.7 | 2.5 | 3.5 |
| 六丝钝尾鰕虎鱼 | 4.1 | 1.1 | 3.6 | 1.6 |
| 红狼牙鰕虎鱼 | 26.3 | 28.2 | — | — |
| 孔鰕虎鱼 | 11.6 | 10.5 | — | — |
| 绿鳍鱼 | 9.1 | 2.0 | — | — |
| 葛氏长臂虾 | 4.7 | 0.8 | — | — |
| 脊尾白虾 | 3.1 | 2.2 | — | — |
| 六指马鲅 | — | — | 7.2 | 6.4 |
| 口虾蛄 | — | — | 5.3 | 4.6 |
| 优势种合计 | 79.8 | 74.1 | 63.9 | 69.2 |
| 头足类 | 0.2 | 0.8 | 1.4 | 1.8 |
| 甲壳类 | 33.4 | 26.9 | 42.0 | 37.8 |
| 鱼类 | 66.4 | 72.3 | 56.6 | 60.4 |
春季10种优势种占总个体数量的79.8%和总渔获生物重量的74.1%, 秋季7种优势种的占比分别为63.9%和69.2%。整体上, 春季优势度较秋季更为集中(图 2, 表 1)。
 |
| 图 2 闽东近海春、秋季游泳动物群落的生物个体数量(a)和重量(b)优势度曲线 Fig. 2 Accumulative percentage curves of individuals (a) and weight (b) of the nekton in the eastern coastal waters of Fujian Province |
基于生物个体数组成分析, 春季, 整个调查海域的生物多样性指数(H′)为2.69, 均匀度指数(J′)为0.67;各站位H′和J′值范围分别为1.37—2.88和0.34—0.72, 均值分别为2.33和0.58。秋季, 整个海域H′值为2.75, J′值为0.67;各站位H′和J′范围为1.58—2.83和0.39—0.69, 均值分别为2.38和0.58 (图 3)。
 |
| 图 3 闽东近海春、秋季游泳动物群落的生物多样性指数(H′)与均匀度指数(J′)的季节性差异 Fig. 3 Seasonal variation of biodiversity index (H′) and evenness index (J′) of the nekton community in the eastern coastal waters of Fujian Province |
基于生物重量组成分析, 春季, 整个调查海域的生物多样性指数(H′)为2.76、均匀度指数(J′)为0.69;各站位H′和J′值分别为1.80—2.93和0.45—0.73, 均值分别为2.50和0.62。秋季, 整个海域H′值为2.96, J′值为0.72;各站位H′和J′范围为2.04—3.02和0.50—0.74, 均值分别为2.61和0.64 (图 3)。
整体上, 基于生物重量的H′和J′值高于基于个体数的计算值; 两种计算情境下, H′和J′值在秋季均高于春季。
2.4.2 地理分布差异各站位的生物多样性指数(H′)和均匀性指数(J′)如图 4所示。
 |
| 图 4 闽东近海游泳动物群落的H′和J′的时空分布 Fig. 4 Spatio-temporal distribution of the H′ and J′ of the nekton community in the eastern coastal waters of Fujian Province 注: a和b:基于生物个体数分析; c和d:基于生物重量分析 |
以各站基于个体数的H′和J′分布为例, 春季, S2站位的H′最高(2.88), S5的H′最低(1.37)。全部站位中, S2—S3、S14和S20—S21的生物多样性水平相对较高(H′ > 2.50), 主要分布于星仔岛东北以及三沙湾西洋岛附近海域。秋季, S2的H′最高(12.83), S10的 H′最低(1.58), 高值站位主要分散于26.5°N以北水域(图 4a)。
春、秋两季的生物多样性最高值均位于闽浙交界的星仔岛东北海域(S2)。另外, 春季, 五个站位(S3、S10、S14、S25和S28)的H′明显高于秋季(ΔH′ > 0.2);秋季, 七个站位(S5—S7、S11、S13、S17和S26)的H′明显高于春季。
均匀性指数J′的地理分布与生物多样性指数H′的变化趋势基本一致(图 4b)。基于生物重量的H′和J′与基于个体数的H′和J′的地理分布趋势基本一致(图 4c, d)。
2.5 单位捕捞努力量渔获量(CPUE)春季, 各站位游泳动物的重量CPUE为11.4—29.2kg/net (均值24.4kg/net), 个体数CPUE为1629—3731ind/net (均值2485ind/net)。其中, 鱼类的平均CPUE为17.64kg/net (72.3%)和1649ind/net (66.4%), 甲壳类为6.55kg/net (26.9%)和829ind/net (33.4%), 头足类为0.20kg/net (0.8%)和7ind/net (0.2%;附录)。十种优势种的生物重量和个体数量CPUE分别占总CPUE的74.1%和79.8%。整体上, 中部水域的个体数量CPUE高于南部和北部, 由近岸向外海呈降低趋势, 在福宁湾(S13)、三沙湾(S17—S18)和星仔岛附近海域(S5)高于3500ind/net, 但重量CPUE分布无明显趋势特征(图 5a, c)。
 |
| 图 5 闽东近海春、秋季游泳动物CPUE的时空分布 Fig. 5 Spatio-temporal distribution in CPUE of the nekton in the eastern coastal waters of Fujian Province 注: a和b分别为春、秋季的个体数CPUE; c和d分别为春、秋季重量CPUE |
秋季, 各站位游泳动物的重量CPUE为8.3—36.3kg/net (均值21.9kg/net); 个体数CPUE为531—2593ind/net (1621ind/net)。其中, 鱼类的平均CPUE分别为13.2kg/net (60.4%)和916ind/net (56.6%), 甲壳类分别为8.28kg/net (37.8%)和680ind/net (42.0%), 头足类分别为0.40kg/net (1.8%)和23ind/net (1.4%;附录)。7个优势种的重量和个体数CPUE分别占总CPUE的69.2%和63.9%。个体数CPUE高值出现在北部的晴川湾、星仔岛(S5)以及南部的闽江口海域(S24—S28), 低值出现在中部的福宁湾、三沙湾及其外海站位(S12—S16;图 5b)。重量CPUE整体上呈南、北部水域高于中部水域和由近岸向外海呈降低趋势, 高值出现在马祖列岛以南(S22)、闽江口东南(S27—S28)以及星仔岛(S3, S5—S9)附近海域(图 5d)。
整体上, 春季各站位生物的重量CPUE与秋季相近, 但其个体数CPUE显著高于秋季。
2.6 资源密度春季, 各站位游泳动物的资源密度为436.9— 1673.2kg/km2 (均值1013.7kg/km2)和5.5×104— 15.7×104ind/km2 (10.2×104ind/km2)。其中, 鱼类的资源密度为756.6kg/km2 (占总资源密度74.6%, 下同)和7.1×104ind/km2 (69.2%), 甲壳类为245.1kg/km2 (24.2%)和3.1×104ind/km2 (30.4%), 头足类为12.0kg/km2 (1.2%)和390ind/km2 (0.4%)。而10种优势物种的资源密度为749.7kg/km2 (73.9%)和8.2×104ind/km2 (80.3%;表 2, 附录)。
| 资源密度 | 调查海域 | 文献来源 | |
| 鱼类 | 全部类群 | ||
| 春: 290.66; 0.8×104 | 春: 366.66; 0.9×104 | 东山湾 | 林龙山等, 2012 |
| 秋: 309.69; 0.6×104 | 秋: 390.27; 1.1×104 | ||
| 春: 327.57; 3.4×105 | 春: 778.24 (扫海法) | 闽江口/九龙江口 | 黄良敏, 2011 |
| 秋: 932.48; 9.9×105 | 秋: 1261.39 | ||
| 春: 553.60 (Δ-分布模型) | |||
| 秋: 2569.11 | |||
| 春: 471.53; 4.1×104 | 九龙江口 | 杜建国等, 2012 | |
| 秋: 143.97; 0.4×104 | |||
| 春: 756.6; 7.1×104 | 春: 1013.7; 10.2×104 | 闽东近海 | 本调查 |
| 秋: 565.7; 4.0×104 | 秋: 899.0; 6.6×104 | ||
| 注:资源密度单位:重量kg/km2; 数量或尾数ind/km2 | |||
|
秋季, 各站位游泳动物的资源密度334.1— 1485.3kg/km2 (899.0kg/km2)和2.2×104—10.5×104ind/km2 (6.6×104ind/km2)。其中, 鱼类的资源密度为565.7kg/km2 (62.9%)和4.0×104ind/km2 (59.8%), 甲壳类为309.7kg/km2 (34.5%)和2.5×104ind/km2 (38.1%), 头足类为23.6kg/km2 (2.6%)和1368ind/km2 (2.1%)。而7种优势物种的资源密度为610.4kg/km2 (67.7%)和4.2×104 ind/km2 (63.3%)。
各站位游泳动物的资源密度的地理分布与其CPUE的地理分布特征类似。
3 讨论本研究中, 春季和秋季调查共出现游泳动物70种, 生物种类数低于九龙江口等福建近海河口区的种类数, 与星仔岛、三沙湾等海域游泳动物种类数水平相近(黄良敏, 2011;沈长春, 2011;张静等, 2013;顾洪静, 2014;张春草, 2017)。河口区由于咸淡水交汇, 陆源营养盐输入丰富, 饵料基础条件优越, 游泳动物群落结构更复杂多样。特别是众多鲤形目鱼类和河口性鱼类喜欢聚集在河口区栖息或洄游, 提高了河口区的物种多样性。而本次的近海调查水域中河口区不多, 河口性鱼类出现较少, 可能导致游泳动物种类偏少。
游泳动物的资源密度呈由近岸至外海逐渐降低的趋势。本次调查共出现鱼类47种、甲壳类18种、头足类5种, 鱼类是主要的生物类群, 其群落组成、个体数或重量资源密度均超过游泳动物总量的50%。这一结果与南麂岛、七星列岛和东山湾等水域的调查结果类似(林龙山等, 2012;晁文春, 2013;何贤保等, 2013;张春草, 2017;叶孙忠等, 2018)。游泳动物群落以鱼类为主的现象可能与调查海域的营养条件有关, 鱼类食性更为复杂, 近海丰富的碎屑有机质、藻类及其他饵料生物可满足不同食性鱼类的摄食生长和繁衍生息。
春季、秋季调查中分别有10种、7种优势种, 优势种存在一定程度的季节性更替, 这与其它相关研究结论相吻合(黄良敏, 2011;张春草, 2017)。优势种存在季节性差异可能与游泳动物栖息、摄食与洄游习性等有关。例如, 六指马鲅是秋季特有的优势种, 未在春季出现。六指马鲅属底栖型鱼类, 夏季在河口区繁育, 秋季则常常向近、外海集群洄游索饵(何贤保等, 2013;张春草, 2017)。葛氏长臂虾、脊尾白虾、孔鰕虎鱼、红狼牙鰕虎鱼和绿鳍鱼为春季优势种。其中, 红狼牙鰕虎鱼在春季渔获中优势度最高。它们同属小型底层生物, 栖息于近岸水域, 主要摄食有机碎屑、小型无脊椎动物等(顾洪静, 2014)。春季, 在沿岸流和温度升高的作用下, 近岸饵料丰度升高, 为众多小型底栖型鱼虾类的产卵、育幼提供了良好的饵料条件。而龙头鱼和三疣梭子蟹是春、秋季的共有优势种, 其秋季资源量显著高于春季, 可能两者均在闽东近海水域生殖和索饵洄游。另外, 口虾蛄是春季的重要种, 同时是秋季优势种, 分布水域较广。本研究的调查水域中的游泳动物群落的生物多样性指数整体上高于东山湾(林龙山等, 2012;叶孙忠等, 2018)和三沙湾(沈长春, 2011), 但低于闽江口、九龙江口及七星列岛海域(黄良敏, 2011;顾洪静, 2014;张春草, 2017), 与台湾海峡(林龙山等, 2005)的生物多样性出于同一水平上。
春季各站位的平均重量CPUE与秋季相近, 但其平均个体数CPUE显著高于秋季, 可能与该海域生物类群的生态类型组成相关。调查中出现的物种多为中小型底栖动物, 在闽东近海进行产卵和索饵育幼。春季(5—8月)是主要繁殖期, 补充群体增多, 幼体比例高。此外, 春季调查中游泳动物资源密度高于东山湾、闽江口和九龙江口; 秋季资源密度高于东山湾, 但低于闽江口和九龙江口的资源密度(黄良敏, 2011;杜建国等, 2012;林龙山等; 2012)。游泳动物的资源密度分布受水陆交互作用、饵料基础、捕捞强度、栖息地功能、鱼类洄游, 甚至保护区的分布等诸多因素影响。秋季, 闽江口的高资源密度可能与河口区良好的饵料基础相关, 该结论与本研究中资源密度分布结论基本一致。另外, 本研究中春季游泳动物资源密度较高可能反映了由于近年来伏季休渔等资源养护措施的积极开展与实施、近海渔业资源得到一定程度恢复这一现象。
4 结论鱼类是闽东近海主要的游泳动物类群, 其种类数和资源量明显高于甲壳类和头足类; 游泳动物群落的优势种存在一定程度季节性更替现象, 春季(10种)多于秋季(7种), 两季共有优势5种; 近岸水域的资源密度相对高于外海水域, 可能与这些水域具有良好的饵料基础有关; 与福建近海其它水域研究结果相比, 闽东近海水域出现的游泳动物种类略低, 但其生物多样性、均匀度水平和平均资源密度均处于较高水平。随着人类对近海的开发利用进一步加剧, 近海渔业资源面临进一步的威胁。因此, 需要通过强化用海活动监督与环境监测, 合理规划、开发利用和保护河口、岛礁、浅海等功能区, 保护鱼类“三场一通道”, 改善渔业环境质量与功能; 或通过海洋牧场建设或其他资源增殖措施开展渔业资源管理和养护, 以保证近海渔业资源可持续发展和利用。
叶孙忠, 罗冬莲, 蔡建堤, 等. 2018. 东山湾渔业生物群落结构及生物量分布特征. 渔业研究, 40(5): 358-365 |
刘勇, 李圣法, 程家骅. 2006. 东海、黄海鱼类群落结构的季节变化研究. 海洋学报, 28(4): 108-114 |
杜建国, 刘正华, 余兴光, 等. 2012. 九龙江口鱼类多样性和营养级分析. 热带海洋学报, 31(6): 76-82 |
何贤保, 章飞军, 林利, 等. 2013. 南麂列岛岛礁区域鱼类种类组成和数量分布. 海洋与湖沼, 44(2): 453-460 |
沈长春. 2011. 福建三沙湾鱼类群落组成特征及其多样性. 海洋渔业, 33(3): 258-264 |
张静, 徐思嘉, 李渊, 等. 2013. 东山湾及其邻近海域鱼类群落结构分析. 中国海洋大学学报, 43(4): 51-57 |
张会军, 2013.福建九龙江河口区鱼类资源现状研究.厦门: 集美大学硕士学位论文
|
张春草, 2017.七星列岛海域游泳动物群落特征及多样性研究.舟山: 浙江海洋大学硕士学位论文
|
林龙山, 郑元甲, 马春艳. 2005. 台湾海峡夏秋季游泳动物资源分布及群落结构. 应用生态学报, 16(10): 1948-1951 |
林龙山, 赵贵根, 李渊, 等. 2012. 东山湾及其邻近海域游泳动物多样性的分析. 水生生物学报, 36(6): 1119-1127 |
顾洪静, 2014.福建九龙江口水域鱼类群落及其资源的研究.厦门: 集美大学硕士学位论文
|
晁文春, 2013.南麂列岛附近海域甲壳类群落结构与生物多样性分析.舟山: 浙江海洋学院硕士学位论文
|
黄良敏, 2011.闽江口和九龙江口及其邻近海域渔业资源现状与鱼类多样性.青岛: 中国海洋大学博士学位论文, 18-20
|
戴泉水, 卢振彬, 戴天元, 等. 2005. 台湾海峡及其邻近海域游泳生物种类组成和资源现状. 水产学报, 29(2): 205-210 |
Pianka E R, 1971. Ecology of the Agamid Lizard Amphibolurus isolepis in Western Australia. Copeia, 1971(3): 527-536 |