 2022, Vol. 53
2022, Vol. 53中国海洋湖沼学会主办。
文章信息
- 马新, 徐金涛, 杨雯, 李微微, 谭法启, 王众, 李欣阳. 2022.
- MA Xin, XU Jin-Tao, YANG Wen, LI Wei-Wei, TAN Fa-Qi, WANG Zhong, LI Xin-Yang. 2022.
- 2021年秦皇岛近岸典型海域浮游植物群落结构的时空变化特征
- SPATIO-TEMPORAL DISTRIBUTION OF PHYTOPLANKTON COMMUNITY IN TYPICAL COASTAL WATERS OF QINHUANGDAO IN 2021
- 海洋与湖沼, 53(6): 1415-1429
- Oceanologia et Limnologia Sinica, 53(6): 1415-1429.
- http://dx.doi.org/10.11693/hyhz20220200035
文章历史
-
收稿日期:2022-02-18
收修改稿日期:2022-04-10




浮游植物是海洋生态系统初级生产力的主要贡献者, 作为食物链的基础环节, 其种群变动和群落结构演替直接影响整个海洋食物链的物质循环和能量流动(Suikkanen et al, 2007; 许歆, 2017)。浮游植物群落可以灵敏而迅速地反映环境的变化, 能够在一定程度上反映海洋的生态环境状态(Charlon et al, 1987; 吴玉霖等, 2004; Padisák et al, 2006; 刘素娟等, 2007)。此外, 浮游植物群落结构的研究能够提高对监测海域长期变化规律的认识, 为预测海洋生态系统结构和功能的变化提供理论依据和数据基础(栾青杉等, 2018)。
秦皇岛位于河北省东北部, 地处北渤海的温带海域, 与渤海海峡相对, 是辽东湾和渤海中部及渤海湾进行物质和能量交换的重要通道(赵相伟等, 2020)。沿岸有洋河、戴河、新河、汤河和新开河等自然河流和多个排污口(Dong et al, 2021), 近年来, 随着经济社会的高速发展, 秦皇岛海域生态环境受到陆源径流及人类活动的强烈干扰, 赤潮时有发生。据统计, 2000~ 2020年在秦皇岛海域共记录赤潮63次(河北省海洋局, 2000~2018; 河北省自然资源厅, 2019-2020; 张万磊等, 2020), 而2011~2019年的10年间是赤潮发生最为集中的时期, 共发生赤潮42次, 甚至出现了一起由链状亚历山大藻(Alexandrium catenella)赤潮引发贻贝体内麻痹性贝毒超标, 人食用后引起的中毒事件(于仁成等, 2016; Yu et al, 2021), 引发各级政府和社会公众的高度关注。其中, 发生赤潮最频繁的两个海域分别是金梦海湾(Jinmeng Bay, 简称JMB)附近海域和东山浴场(Dongshan Beach, 简称DSB)附近海域。
金梦海湾和东山浴场分别是秦皇岛市西部和东部比较有代表性的海滨浴场, 周围均有居民区、餐饮酒店和旅游景点等, 金梦海湾紧邻大、小汤河入海口以及东北大学生活污水排污口和电子厂工业污水排污口, 东山浴场紧邻秦皇岛港、新开河入海口以及热电里小区生活污水排污口和热电厂工业污水排污口。2011~2019年的42次赤潮中, 有28次赤潮的发生涉及这两个海域, 年均发生赤潮3.1次; 2015年和2016年是发生赤潮次数最多的年份, 共11次; 共有22种赤潮生物形成赤潮, 形成赤潮次数最多的藻种为夜光藻(Noctiluca scintillans, 10次)、锥状斯克里普藻(Scrippsiella trochoidea, 7次)、赤潮异弯藻(Heterosigma akashiwo, 4次)和抑食金球藻(Aureococcus anophagefferens, 4次)等。
然而, 对该海域浮游植物群落结构的研究较少, 相关调查多集中在2015年以前(张建乐等, 2007; 慕建东等, 2015; 于湖洋等, 2016; 许歆, 2017), 近年来尚无对该海域浮游植物群落的研究报道。本研究基于2021年4~10月秦皇岛近岸海域网采浮游植物的逐月调查结果, 讨论了该海域网采浮游植物的种类组成、优势种、丰度、多样性等特征及其与环境因子的关系, 分析了金梦海湾和东山浴场两个典型海域海水水质和浮游植物群落结构的异同, 以期为该海域浮游植物群落状况研究和赤潮的预警监测提供数据依据。
1 材料与方法 1.1 站位布设本研究在秦皇岛近岸海域设置12个调查站位, 于2021年4~10月开展了7个航次浮游植物和环境因子的逐月调查。调查范围及站位分布见图 1, 其中, S11~S16站位位于金梦海湾附近海域, S21~S26站位位于东山浴场附近海域。
 |
| 图 1 调查站位示意图 Fig. 1 Schematic drawing of stations |
浮游植物调查: 参照《海洋调查规范第6部分》(GB/T 12763.1-2007)进行浮游植物采样, 采用浅水Ⅲ型浮游生物网(网口直径37 cm, 网口面积0.1 m2, 网径76 μm)自水体底层至表层垂直拖网1次, 收集浮游植物样品后立即加体积比为5%的甲醛固定。带回实验室后, 将浮游植物样品充分混匀, 取0.1~0.5 mL用Olympus BX53生物显微镜进行鉴定和计数。
水环境调查: 水样的采集、现场处理和分析测定均按《海洋监测规范》(GB 17378.4-2007)和《海洋调查规范》(GB/T 12763.4-2007)中规定的方法执行。涉及的水质参数有水温、盐度、pH值、溶解氧、化学需氧量COD、硝酸盐、亚硝酸盐、氨盐、活性磷酸盐和活性硅酸盐。
1.3 数据处理与分析 1.3.1 环境因子选用富营养化指数(E)来评价海水富营养化程度, 计算公式为
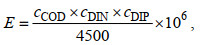 (1)
(1)式中, E为富营养化指数, 无量纲; cCOD为化学需氧量浓度, mg/L: cDIN为无机氮浓度, mg/L; cDIP为无机磷浓度, 在本文中以磷酸盐浓度代替, mg/L。一般认为E≥1为富营养化, 其中1≤E≤3时为轻度富营养化, 3 < E≤9时为中度富营养化, E > 9为重度富营养化(王燕等, 2021)。
1.3.2 优势种浮游植物优势种以优势度指数Y判断, 一般将Y≥0.02的种类定义为优势种(Dufrene et al, 1997), 优势度Y值计算公式为
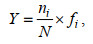 (2)
(2)式中, N为调查站位所有浮游植物出现的总个体数; ni为第i种浮游植物的个体数; fi为第i种浮游植物在各站位的出现频率。
1.3.3 生物多样性用Shannon-Wiener多样性指数(H′)(Shannon et al, 1949)和Pielou均匀度(J)(Pielou, 1969)进行浮游植物群落多样性分析。
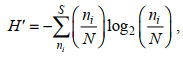 (3)
(3)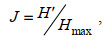 (4)
(4)式中, S为群落中的总物种数; Hmax表示最大物种多样性指数log2S。
1.3.4 群落结构及环境因子分析主要由Primer 6.0软件完成。采用CLUSTER模块对浮游植物群落进行聚类分析, 分析不同月份间浮游植物群落结构的差异情况, 并对其进行One-way ANOSIM检验; 采用MDS模块形成的二维平面图来解释不同月份间的群落结构相似性, 其可信程度用胁强系数(Stress)来衡量, 一般当Stress < 0.2时, 认为该图的结果是可信的, 有一定的解释意义(Field et al, 1982); 采用BIOENV模块分析得出对浮游植物群落相关性最密切的环境因子组合, 并对其进行RELATE检验。
1.3.5 其他采用surfer 11.0软件绘制站位分布图和浮游植物丰度平面分布图, 其余作图及数据的统计分析由Origin 8.5软件完成。
2 结果与分析 2.1 环境因子金梦海湾、东山浴场和秦皇岛近岸海域(指整个研究海域, 下同)7个月份的水质参数均值结果见图 2。从研究海域整体看, 海水水温有明显的季节变化, 4月最低(11.9 ℃), 8月最高(26.8 ℃); 4~6月盐度较高(31.95~32.53), 7~10月盐度下降, 稳定在30左右; 溶解氧、pH值均符合第一类海水水质标准; COD除8月和9月超出2 mg/L, 属于第二类海水水质外, 其余月份符合第一类海水水质标准; 10月无机氮浓度高于0.2 mg/L, 属于第二类海水水质, 其余月份满足第一类海水水质标准; 活性磷酸盐浓度均符合第一类海水水质标准, 但呈现先降低后升高的趋势, 6~8月浓度较低, 氮磷比极高(79.65~149.38); 活性硅酸盐浓度全年均处于较高水平, 尤其是8月份, 达1.09 mg/L。7个航次富营养化指数范围为0.04~1.04, 除10月份略超富营养化水平外, 该海域绝大部分时间水质远未达到富营养化水平。总体而言, 研究海域2021年水质状况良好。
 |
| 图 2 金梦海湾、东山浴场和秦皇岛近岸海域水质参数的月际变化 Fig. 2 Intermonthly variation of water quality parameters in the Jinmeng Bay, Dongshan Beach, and Qinhuangdao costal area |
比较来看, 金梦海湾和东山浴场两个典型海域水温的变化趋势比较一致, 最低水温均出现在4月(金梦海湾附近海域和东山浴场附近海域分别为12.5和11.4 ℃, 顺序下同), 最高水温均出现在8月(分别为27.0和26.5 ℃)。4~8月金梦海湾附近海域较东山浴场附近海域水温偏高近1.0 ℃, 8~9月偏低约0.6 ℃。综合其他7项水质参数结果, 金梦海湾附近海域各项理化参数的变化范围均明显大于东山浴场附近海域, 尤其是盐度、pH值、溶解氧、硅酸盐浓度和磷酸盐浓度, 而且这些相对剧烈的波动多出现在7~9月。在此期间, 金梦海湾附近海域的海水盐度呈断崖式下降, pH值、溶解氧、COD和硅酸盐浓度则是先升高后下降, 而9月磷酸盐浓度则显著高于东山浴场附近海域。从营养盐浓度来看, 东山浴场附近海域磷酸盐浓度低于金梦海湾附近海域, 5~7月部分站位存在磷限制, 但两个海域均没有氮限制或硅限制现象。
2.2 浮游植物 2.2.1 种类组成研究海域在2021年4~10月共鉴定出浮游植物3门45属92种(见附录1), 其中, 硅藻门32属67种, 占总种类数的72.8%; 甲藻门12属24种, 占26.1%; 金藻门1属1种, 占1.1%。硅藻是绝对的优势类群, 其中又以角毛藻属(Chaetoceros)的种类最多, 达20种, 其次为菱形藻属(Nitzschia)和圆筛藻属(Coscinodiscus), 分别为5种和4种。甲藻门中以原多甲藻属(Protoperidinium)的种类最多, 有8种, 其次为角藻属(Ceratium), 有4种。9月浮游植物种类数最多, 共3门50种, 其中, 硅藻门36种, 甲藻门13种, 金藻门1种; 7月和8月浮游植物种类数次之, 均为46种, 5月浮游植物种类数最少, 共2门21种(图 3)。
 |
| 图 3 金梦海湾、东山浴场和秦皇岛近岸海域浮游植物种类组成的月际变化 Fig. 3 Intermonthly variation of species composition in the Jinmeng Bay, Dongshan Beach, and Qinhuangdao coastal area |
比较来看, 金梦海湾附近海域共鉴定浮游植物3门42属83种, 其中, 硅藻门29属60种, 甲藻门12属22种, 金藻门1属1种; 东山浴场附近海域共鉴定浮游植物39属82种, 其中硅藻门27属59种, 甲藻门11属22种, 金藻门1属1种。两个海域浮游植物种类组成差别不大, 但东山浴场附近海域的种类数普遍高于金梦海湾附近海域的种类数(图 3)。
2.2.2 优势种种类及其变化研究海域浮游植物各月份出现的优势种及优势度Y值见表 1。不同月份浮游植物优势种的种类和数量差异很大, 9月优势种数量最多, 共10属12种, 除裸甲藻(Gymnodinium sp.)外, 其余均为硅藻, 其中优势地位最明显的是劳氏角毛藻(Chaet. lorenzianus); 7月优势种数量次之, 共5属8种, 角毛藻属和角藻属各3种, 中肋骨条藻(Skeletonema costatum)和三角角藻(C. tripos)优势度最高; 6月有6属6种, 星脐圆筛藻(Cos. asteromphalus)占绝对优势, 夜光藻次之; 10月有4属5种, 星脐圆筛藻和尖刺伪菱形藻(Pseudo-nitzschia pungens)占优势地位。4月和5月各有4种优势种, 但4月以扭链角毛藻(Chaet. tortissimus)占绝对优势, 5月星脐圆筛藻、梭角藻(C. fusus)占绝对优势, 其次为夜光藻。8月优势种数量最少, 只有2属2种, 中肋骨条藻具有非常高的优势地位, 其丰度占浮游植物总丰度的90.9%, 其次为窄隙角毛藻(Chaet. affinis)。未出现7个航次共有的优势种。
| 优势种 | 优势度Y值 | ||||||||
| 序号 | 中文名 | 拉丁名 | 4月 | 5月 | 6月 | 7月 | 8月 | 9月 | 10月 |
| 1 | 派格棍形藻 | Bacillaria paxillifera | 0.07 | ||||||
| 2 | 窄隙角毛藻 | Chaetoceros affinis | 0.04 | 0.07 | 0.03 | ||||
| 3 | 旋链角毛藻 | Chaetoceros curvisetus | 0.04 | ||||||
| 4 | 双孢角毛藻 | Chaetoceros didymus | 0.02 | ||||||
| 5 | 劳氏角毛藻 | Chaetoceros lorenzianus | 0.23 | ||||||
| 6 | 角毛藻 | Chaetoceros sp. | 0.03 | ||||||
| 7 | 扭链角毛藻 | Chaetoceros tortissimus | 0.78 | ||||||
| 8 | 星脐圆筛藻 | Coscinodiscus asteromphalus | 0.02 | 0.35 | 0.51 | 0.08 | 0.35 | ||
| 9 | 威利圆筛藻 | Coscinodiscus wailesii | 0.04 | ||||||
| 10 | 筛链藻 | Coscinosira sp. | 0.03 | ||||||
| 11 | 短角弯角藻 | Eucampia zoodiacus | 0.02 | ||||||
| 12 | 斯氏几内亚藻 | Guinardia striata | 0.10 | ||||||
| 13 | 丹麦细柱藻 | Leptocylindrus danicus | 0.03 | ||||||
| 14 | 微小细柱藻 | Leptocylindrus minimus | 0.02 | ||||||
| 15 | 柔弱伪菱形藻 | Pseudo-nitzschia delicatissima | 0.04 | 0.11 | |||||
| 16 | 尖刺伪菱形藻 | Pseudo-nitzschia pungens | 0.04 | 0.26 | |||||
| 17 | 中肋骨条藻 | Skeletonema costatum | 0.19 | 0.91 | 0.10 | ||||
| 18 | 掌状冠盖藻 | Stephanopyxis palmeriana | 0.05 | ||||||
| 19 | 佛氏海线藻 | Thalassionema frauenfeldii | 0.03 | ||||||
| 20 | 圆海链藻 | Thalassiostra rotula | 0.04 | ||||||
| 21 | 亚历山大藻 | Alexandrium sp. | 0.09 | ||||||
| 22 | 叉角藻 | Ceratium furca | 0.08 | ||||||
| 23 | 梭角藻 | Ceratium fusus | 0.05 | 0.34 | 0.07 | 0.04 | |||
| 24 | 三角角藻 | Ceratium tripos | 0.13 | ||||||
| 25 | 裸甲藻 | Gymnodinium sp. | 0.04 | ||||||
| 26 | 夜光藻 | Noctiluca scintillans | 0.08 | 0.17 | 0.15 | ||||
比较来看, 金梦海湾附近海域有24个优势种, 9月优势种数量最多, 共8属10种, 5月最少, 共3属3种; 星脐圆筛藻在5个月份的调查中成为优势种, 其次为梭角藻和夜光藻, 均为4次。东山浴场附近海域也有24个优势种, 9月优势种数量最多, 共8属10种, 8月最少, 仅有1属1种; 星脐圆筛藻在5个月份的调查中成为优势种, 其次为中肋骨条藻、夜光藻和梭角藻, 均为3次。
2.2.3 丰度及其平面分布金梦海湾、东山浴场和秦皇岛近岸海域浮游植物丰度的月际变化见图 4。研究海域浮游植物丰度范围为(1.09~101.49)×106 cells/m3, 平均值为1.67×107 cells/m3, 最高值出现在8月, 最低值出现在6月。金梦海湾附近海域浮游植物丰度范围为(0.51~37.99)×106 cells/m3, 平均值为7.34×106 cells/m3, 最高值出现在8月, 最低值出现在4月。东山浴场附近海域浮游植物丰度范围为(0.93~164.98)×106 cells/m3, 平均值为2.60×107 cells/m3, 最高值出现在8月, 最低值出现在6月。比较来看, 东山浴场附近海域除4月浮游植物丰度显著高于金梦海湾附近海域(P < 0.05)外, 其他月份的浮游植物丰度与金梦海湾附近海域无显著差异(P > 0.05)。
 |
| 图 4 金梦海湾、东山浴场和秦皇岛近岸海域浮游植物丰度的月际变化 Fig. 4 Intermonthly variation of phytoplankton levels in the Jinmeng Bay, Dongshan Beach, and Qinhuangdao coastal area |
研究海域浮游植物丰度的平面分布见图 5。4月, 浮游植物丰度高值区在以S21为代表的东山浴场附近海域(平均值为3.39×106 cells/m3), 金梦海湾附近浮游植物丰度很低(平均值为5.13×105 cells/m3); 5月浮游植物丰度分布较均匀, 呈现近岸高, 远岸低的特点, 与4月相比, 东山浴场附近海域丰度下降, 金梦海湾附近海域丰度升高; 6月, 浮游植物丰度为调查期间的最低值, 呈块状分布, 两个浴场之间的海域浮游植物丰度最低; 7月浮游植物丰度有所回升, 均值为2.23×106 cells/m3, 并呈现西部海域低, 东部海域高的趋势; 8月整个海域的浮游植物丰度达到最高峰, 以S23 (3.16×108 cells/m3)和S24 (3.89×108 cells/m3)为中心的东山浴场海域的平均丰度(1.65×108 cells/m3)显著高于金梦海湾海域的平均丰度(3.80×107 cells/m3), 中肋骨条藻贡献了90%以上的丰度值, 其他优势种类也多以角毛藻为主; 9月浮游植物丰度大幅下降, 并在研究海域中部出现以S13为中心的高值区, 并以此为中心向四周扩展, 丰度逐渐下降; 10月丰度与9月差别不大, 但高值区向东南部海域扩展, 西南部海域和东北部海域的浮游植物丰度极低。
 |
| 图 5 2021年4~10月秦皇岛附近海域浮游植物丰度的平面分布图 Fig. 5 Horizontal distribution of phytoplankton abundance from April to October in 2021 |
另外, 金梦海湾和东山浴场附近海域硅甲藻的丰度比值差别明显(图 6)。除5月和7月金梦海湾附近海域的硅甲藻丰度比值小于1, 即甲藻丰度高于硅藻外, 其他月份硅藻丰度均高于甲藻。5月金梦海湾附近海域的甲藻优势种为梭角藻和夜光藻, 而7月为三角角藻、叉角藻和梭角藻。以8月为界, 金梦海湾附近海域和东山浴场附近海域的硅甲藻丰度比值出现逆转, 即4~8月金梦海湾附近海域浮游植物群落中甲藻丰度占比高于东山浴场附近海域, 而9月和10月则相反。
 |
| 图 6 2021年4~10月金梦海湾、东山浴场附近海域硅甲藻丰度比 Fig. 6 The abundance ratio of diatoms to dinoflagellates in the coastal areas of Jinmeng Bay and Dongshan Beach from April to October in 2021 |
研究海域浮游植物群落的多样性指数变化范围为1.19~3.43, 平均值为2.09, 最高值出现在9月, 最低值出现在8月; 金梦海湾附近海域浮游植物群落的多样性指数变化范围为1.49~3.24, 平均值为2.18, 最高值出现在9月, 最低值出现在5月; 东山浴场浮游植物群落的多样性指数变化范围为0.49~3.62, 平均值为2.00, 最高值出现在9月, 最低值出现在8月。
研究海域浮游植物群落的均匀度变化范围为0.26~0.74, 平均为0.54, 最高值出现在9月, 最低值出现在8月; 金梦海湾附近海域浮游植物群落的均匀度变化范围为0.41~0.74, 平均值为0.58, 最高值出现在9月, 最低值出现在8月; 东山浴场附近海域浮游植物群落的均匀度变化范围为0.11~0.74, 平均值为0.51, 最高值出现在9月, 最低值出现在8月。
2.3 群落结构及环境因子分析 2.3.1 聚类分析和MDS标序浮游植物样品经实验室定量分析后, 建立丰度原始矩阵。为减少稀有种对浮游植物群落结构的干扰, 筛选出全年出现频率大于15%的种类, 共22属37种, 将其丰度矩阵进行四次方根转化和Bray-Curtis相似性计算, 经Simprof检验后进行CLUSTER聚类分析和MDS标序, 结果见图 8。以32%相似度为界, 可将调查海域7个月份84个站次的浮游植物群落划分为3组, a组为4、5和6月12个站位的调查结果, b组为9、10月12个站位的调查结果, c组为7、8月12个站位的调查结果。3个组内部的聚类效果各有不同, a组内部的浮游植物群落相似性最高, 其次为b组, c组内部浮游植物群落差异最大。总体来说, 浮游植物的群落组成体现了非常明显的季节特征, 说明浮游植物群落组成随季节变化的分布特征具有良好的稳定性和重现性。二维MDS标序图的结果与CLUSTER聚类分析图一致, Stress值为0.13, 处于可信范围内。
 |
| 图 7 金梦海湾、东山浴场和秦皇岛近岸海域浮游植物多样性指数、均匀度的月际变化 Fig. 7 Intermonthly variation in diversity index H′ and equitability J of phytoplankton in the Jinmeng Bay, Dongshan Beach, and Qinhuangdao coastal area |
 |
| 图 8 秦皇岛近岸海域浮游植物群落的聚类分析图和MDS标序图 Fig. 8 Cluster analysis and MDS of phytoplankton community in Qinhuangdao coastal area |
One-way ANOSIM检验证实了以浮游植物丰度矩阵划分的3个组群之间以及任意2组群之间的差异显著(R=0.915, P=0.001; 成对检验结果见表 2)。
| 组群 | R统计值 | 显著性水平/% |
| a, c | 0.916 | 0.1 |
| a, b | 0.986 | 0.1 |
| c, b | 0.758 | 0.1 |
对该海域浮游植物和水质参数的BIOENV分析结果(表 3)表明, 使环境欧氏距离非相似性矩阵与样品Bray-Curtis非相似性矩阵之间形成最大等级相关的水质参数是水温和盐度的组合, 其等级相关系数为0.513。RELATE检验显示, 浮游植物丰度矩阵与环境变量矩阵的样本统计显著性水平为0.1%, 表明秦皇岛海域所监测的环境因子会影响其浮游植物群落结构。
| 变量 | 相关系数(Spearman) | 环境因子组合 | 变量 | 相关系数(Spearman) | 环境因子组合 |
| 2 | 0.513 | 水温、盐度 | 2 | 0.453 | pH、水温 |
| 3 | 0.506 | pH、水温、盐度 | 1 | 0.446 | 水温 |
| 3 | 0.469 | 水温、氨盐、活性硅酸盐 | 3 | 0.440 | 亚硝酸盐、氨盐、活性硅酸盐 |
| 3 | 0.459 | 水温、亚硝酸盐、活性硅酸盐 | 3 | 0.434 | 溶解氧、氨盐、活性硅酸盐 |
| 1 | 0.458 | 盐度 | 3 | 0.430 | 溶解氧、亚硝酸盐、活性硅酸盐 |
此次调查共鉴定秦皇岛海域浮游植物3门45属92种, 与慕建东等(2015)于2010年5月至9月期间在该海域的调查结果(46属96种)相近, 高于于湖洋等(2016)于2014年4~9月对该海域的调查结果(3门33属70种)。和渤海其他海域调查结果类似(孙萍等, 2008; 于潇等, 2020; 张雪等, 2020), 秦皇岛近岸海域浮游植物种类组成以硅藻为主。
由于秦皇岛海域独特的地理位置, 沿岸入海河流和排污口较多, 个别月份盐度明显偏低, 使其浮游植物以适低盐的近岸种、温带种和广布种为主, 也有少量外海性暖水种和冷水种。26种优势种中有20种硅藻, 6种甲藻, 其中星脐圆筛藻、夜光藻、梭角藻和三角角藻在7个航次均有出现。但从丰度高值看, 适温相对较高的星脐圆筛藻(俞建銮等, 1993)丰度高值出现在10月, 适温较低的夜光藻(曹新宇等, 2021)丰度高值出现在4~6月。与俞建銮等(1993)认为梭角藻往往在8月成为优势种的结论不同, 秦皇岛海域的梭角藻在4~6月成为优势种, 丰度高值出现在5月。三角角藻的丰度高值出现在7月, 该藻随着黑潮和黄海暖流的余脉从渤海海峡输入渤海海域, 属于暖海大洋性的外源性物种(孙军等, 2005; 陈楠生等, 2021)。甲藻优势种多集中出现在4~7月。
3.2 浮游植物丰度秦皇岛近岸海域浮游植物丰度范围为(1.09~ 101.49)×106 cells/m3, 6月最低, 8月最高, 平均值为1.67×107 cells/m3。与历史同期调查结果相比, 2021年秦皇岛近岸海域的浮游植物丰度显著高于2004年(张建乐等, 2007)和2010年(慕建东等, 2015)的浮游植物丰度, 但低于2014年(于湖洋等, 2016)的浮游植物丰度。同一海域不同调查年份间的浮游植物丰度不同可能与研究海域站位布设范围或调查时间不同有关, 如于湖洋等(2016)在秦皇岛的研究海域包含养殖区, 海水营养物质异常丰富, 造成其浮游植物丰度较高, 张建乐等(2007)等在秦皇岛海域的调查时间仅为8月, 慕建东等(2015)在2010年的调查时间和调查海域与本研究最为吻合, 但其浮游植物丰度明显低于本研究, 主要原因是7~8月的浮游植物丰度极低, 可能与当年持续不断发生的赤潮和褐潮灾害有关。东山浴场附近海域的甲藻丰度仅在5月和10月与硅藻丰度相近, 其余时间硅藻均占绝对优势, 而金梦海湾附近海域的甲藻丰度在4~7月与硅藻丰度相近, 7月甚至远高于硅藻丰度, 只在8~10月才表现出明显的硅藻优势。这与历年两个典型海域的甲藻赤潮发生情况相吻合(河北省海洋局, 2000~2018), 东山浴场的甲藻赤潮多为夜光藻赤潮, 发生时间也集中在5~6月, 金梦海湾附近海域发生的甲藻赤潮具有种类多、频率高、时间长的特点, 这可能与该海域春夏季较低的磷酸盐浓度和水动力条件欠佳有关。多项调查(谢琳萍等, 2012; 郭术津等, 2014; 张海波等, 2019)及本研究(图 2)均显示, 渤海海域春夏季多有潜在的磷限制现象出现, 氮磷比较高, 同时受该海域近岸人工岛和进岛路工程的联合作用, 金梦海湾附近海域外海海水与汤河口和近岸的水体交换缓慢(匡翠萍等, 2019)。而在相对稳定且高氮磷比的水体环境中, 更适合有运动能力且更能耐受低磷环境的甲藻的生长, 更易发生甲藻赤潮(Riegman et al, 1996; Egge, 1998)。
3.3 浮游植物群落结构及其与环境因子的关系BIOENV分析结果(表 3)表明, 水温和盐度是影响其浮游植物群落结构的主要环境因子。秦皇岛近岸海域海水水温和盐度呈现明显的季节变化(图 2): 4~6月基本为低温高盐海水; 7~8月为高温低盐海水, 金梦海湾附近海域海水的这一特点更明显; 9~10月水温急剧下降(由24.5 ℃左右降至15.1 ℃左右), 盐度较7、8月略有升高。与水环境月际变化规律相应, 秦皇岛近岸海域浮游植物群落结构也分为3组(图 8), 分别为4~6月的a组, 9~10月的b组和7~8月的c组, 反映在优势种上的更替规律为: a组以广温广盐型和适低温型的藻种, 如扭链角毛藻、星脐圆筛藻、梭角藻和夜光藻为主, b组海水环境变化剧烈, 优势种种类繁杂, c组以广温广盐型的中肋骨条藻和适温较高的各类角毛藻、三角角藻为主。
甲藻优势种多集中出现在4~7月, 其中4~6月的甲藻优势种均为广温广盐种梭角藻和适低温种夜光藻, 7月时, 适低温的夜光藻已不具优势, 取而代之的是高温外海种叉角藻(刘述锡等, 2013)和三角角藻。硅藻为全年的优势类群, 又以8月的中肋骨条藻为甚。该藻虽然只出现在7~9月, 但8月的绝对丰度优势仍使其成为周年优势度最高(Y=0.34)的藻种。中肋骨条藻是一种在全球近岸海域分布极广的广温广盐型的浮游硅藻, 金德祥等(1965)对其进行实验发现最适温盐范围为20~25 ℃和25~30, 而霍文毅等(2001)在对胶州湾一次中肋骨条藻赤潮的研究中发现该藻的最适温盐范围为24~28 ℃和20~30, 发生赤潮时的海水环境多为高温低盐的咸淡混合水。刘东艳等(2002)研究了不同氮磷比对中肋骨条藻生长的影响, 发现氮比磷更易诱发中肋骨条藻赤潮, 即使在高氮磷比情况下, 中肋骨条藻仍能够很好生长。虽然7月~8月的秦皇岛近岸海域海水中的氮磷比较高, 但海水环境(水温范围26.0~26.8 ℃, 盐度范围为29.0~29.6)正符合中肋骨条藻对温盐的最适生长要求, 加之8月海水中高浓度的硅酸盐(图 2)为其生长提供了硅藻所必需的硅元素, 使得中肋骨条藻呈暴发式繁殖。与此相似, 2015年8月在秦皇岛近岸海域曾发生一次中肋骨条藻赤潮。
4 结论(1) 2021年4~10月秦皇岛近岸海域共发现浮游植物3门45属92种, 硅藻门和甲藻门为主要的浮游植物类群; 硅藻门的优势地位明显。浮游植物生态类型以近岸种、温带种和广布种为主, 也有少量外海性暖水种和冷水种。
(2) 不同月份浮游植物优势种的种类和数量差异很大, 9月优势种多样性最高, 8月优势种多样性最低。甲藻优势种多集中出现在4~7月。未出现7个航次共有的优势种。
(3) 浮游植物丰度8月为全年最高峰, 达1.01× 108 cells/m3, 其余月份均在106 cells/m3量级。浮游植物多样性指数和均匀度与优势种多样性保持一致, 9月最高, 8月最低。
(4) 秦皇岛近岸海域浮游植物群落有明显的季节更替现象, 水温和盐度是影响其浮游植物群落结构的主要环境因子。
(5) 金梦海湾和东山浴场两个典型海域的浮游植物种类组成差别不大, 但前者有相对稳定且高氮磷比的水体环境, 更适合甲藻生长, 更容易发生甲藻赤潮。
| 类群 | 种类 | 4月 | 5月 | 6月 | 7月 | 8月 | 9月 | 10月 |
| 硅藻门 | ||||||||
| 短柄曲壳藻Achnanthes brevipes | + | + | + | |||||
| 六幅辐裥藻Actinoptychus senarius | + | |||||||
| 冰河拟星杆藻Asterionellopsis glacialis | + | + | ||||||
| 派格棍形藻Bacillaria paxillifera | + | + | + | |||||
| 透明辐杆藻Bacteriastrum hyalinum | + | + | + | |||||
| 正盒形藻Biddulphia biddulphiana | + | |||||||
| 窄隙角毛藻Chaetoceros affinis | + | + | + | + | + | + | ||
| 卡氏角毛藻Chaetoceros castracanei | + | + | ||||||
| 扁面角毛藻Chaetoceros compressus | + | + | + | |||||
| 深环沟角毛藻Chaetoceros constrictus | + | |||||||
| 发状角毛藻Chaetoceros crinitus | + | |||||||
| 旋链角毛藻Chaetoceros curvisetus | + | + | + | + | ||||
| 柔弱角毛藻Chaetoceros debilis | + | |||||||
| 并基角毛藻Chaetoceros decipiens f. decipiens | + | + | ||||||
| 密连角毛藻Chaetoceros densus | + | + | + | + | + | |||
| 双孢角毛藻Chaetoceros didymus | + | + | + | + | + | |||
| 远距角毛藻Chaetoceros distans | + | |||||||
| 克尼角毛藻Chaetoceros knipowitschii | + | |||||||
| 劳氏角毛藻Chaetoceros lorenzianus | + | + | + | |||||
| 窄面角毛藻Chaetoceros paradoxus | + | + | + | + | ||||
| 拟旋链角毛藻Chaetoceros pseudocurvisetus | + | + | ||||||
| 暹罗角毛藻Chaetoceros siamense | + | + | + | |||||
| 角毛藻Chaetoceros sp. | + | + | + | + | ||||
| 圆柱角毛藻Chaetoceros teres | + | + | + | + | ||||
| 扭链角毛藻Chaetoceros tortissimus | + | + | + | |||||
| 范氏角毛藻Chaetoceros vanheurcki | + | |||||||
| 星脐圆筛藻Coscinodiscus asteromphalus | + | + | + | + | + | + | + | |
| 格氏圆筛藻Coscinodiscus granii | + | + | ||||||
| 琼氏圆筛藻Coscinodiscus jonesianus | + | + | + | + | ||||
| 威利圆筛藻Coscinodiscus wailesii | + | + | + | |||||
| 筛链藻Coscinosira sp. | + | |||||||
| 矮小短棘藻Detonula pumila | + | |||||||
| 布氏双尾藻Ditylum brightwellii | + | + | ||||||
| 唐氏藻Donkinia sp. | + | |||||||
| 短角弯角藻Eucampia zoodiacus | + | + | ||||||
| 脆杆藻Fragilaria sp. | + | |||||||
| 柔弱几内亚藻Guinardia delicatula | + | |||||||
| 薄壁几内亚藻Guinardia flaccida | + | + | ||||||
| 斯氏几内亚藻Guinardia striata | + | + | + | + | ||||
| 中华半管藻Hemiaulus sinensis | + | |||||||
| 丹麦细柱藻Leptocylindrus danicus | + | + | + | + | + | + | ||
| 微小细柱藻Leptocylindrus minimus | + | + | + | + | ||||
| 具槽直链藻Melosira sultana | + | |||||||
| 膜状缪氏藻Meuniera membranacea | + | |||||||
| 舟形藻Navicula sp. | + | + | ||||||
| 新月菱形藻Nitzschia closterium | + | + | + | + | + | + | ||
| 长菱形藻Nitzschia longissima | + | + | ||||||
| 洛氏菱形藻Nitzschia lorenziana | + | + | ||||||
| 琴式菱形藻Nitzschia panduriformis | + | |||||||
| 菱形藻Nitzschia sp. | + | + | + | + | ||||
| 长耳齿状藻Odontella aurita | + | |||||||
| 高齿状藻Odontella regia | + | |||||||
| 羽纹藻Pinnularia sp. | + | + | + | |||||
| 海洋曲舟藻Pleurosigma pelagicum | + | + | ||||||
| 曲舟藻Pleurosigma sp. | + | + | + | |||||
| 柔弱伪菱形藻Pseudo-nitzschia delicatissima | + | + | + | + | ||||
| 尖刺伪菱形藻Pseudo-nitzschia pungens | + | + | + | + | ||||
| 翼根管藻印度变型Rhizosolenia alata f. indica | + | + | + | + | + | |||
| 透明根管藻Rhizosolenia hyalina | + | |||||||
| 刚毛根管藻Rhizosolenia setigera | + | |||||||
| 优美旭氏藻矮小变型Schrö-derella delicatula f. schrö-deri | + | + | + | |||||
| 中肋骨条藻Skeletonema costatum | + | + | + | + | ||||
| 掌状冠盖藻Stephanopyxis palmeriana | + | + | ||||||
| 泰晤士扭鞘藻Streptothece thamesis | + | + | ||||||
| 佛氏海线藻Thalassionema frauenfeldii | + | + | + | + | ||||
| 菱形海线藻Thalassionema nitzschioides | + | + | ||||||
| 圆海链藻Thalassiostra rotula | + | + | ||||||
| 甲藻门 | ||||||||
| 红色赤潮藻Akashiwo sanguinea | + | + | + | |||||
| 亚历山大藻Alexandrium sp. | + | + | + | + | ||||
| 叉角藻Ceratium furca | + | + | + | + | + | |||
| 梭角藻Ceratium fusus | + | + | + | + | + | + | + | |
| 马西利亚角藻Ceratium massiliense | + | + | + | |||||
| 三角角藻Ceratium tripos | + | + | + | + | + | + | + | |
| 渐尖鳍藻Dinophysis acuminata | + | + | + | |||||
| 裸甲藻Gymnodinium sp. | + | + | + | |||||
| 螺旋环沟藻Gyrodinium spirale | + | + | + | + | + | + | ||
| 卡尔藻Karlodinium sp. | + | |||||||
| 夜光藻Noctiluca scintillans | + | + | + | + | + | + | + | |
| 闪光原甲藻Prorocentrum micans | + | |||||||
| 微小原甲藻Prorocentrum minimum | + | + | + | |||||
| 尖叶原甲藻Prorocentrum triestinum | + | + | ||||||
| 双脚原多甲藻Protoperidinium bipes | + | |||||||
| 锥状原多甲藻Protoperidinium conicum | + | |||||||
| 大原多甲藻Protoperidinium grande | + | + | ||||||
| 海洋原多甲藻Protoperidinium oceanicum | + | + | + | |||||
| 灰甲原多甲藻Protoperidinium pellucidum | + | + | ||||||
| 五角原多甲藻Protoperidinium pentagonum | + | |||||||
| 原多甲藻Protoperidinium sp. | + | + | + | + | + | |||
| 斯氏扁甲藻Pyrophacus steinii | + | + | ||||||
| 锥状斯克里普藻Scrippsiella trochoidea | + | + | + | |||||
| 灵巧原多甲藻Protoperidinium venustum | + | |||||||
| 金藻门 | ||||||||
| 小等刺硅鞕藻Dictyocha fibula | + | + | + | + | ||||
| 注: “+”代表该藻种在该月份有出现。 | ||||||||
于仁成, 刘东艳, 2016. 我国近海藻华灾害现状、演变趋势与应对策略. 中国科学院院刊, 31(10): 1167-1174 |
于湖洋, 崔磊, 潘霖, 等, 2016. 秦皇岛海域浮游植物的群落结构特征. 海洋科学, 40(5): 66-75 |
于潇, 卢钰博, 李希磊, 等, 2020. 莱州湾浮游植物时空变化及其与环境因子的关系. 烟台大学学报(自然科学与工程版), 33(1): 63-71 |
王燕, 姚振童, 祝艳君, 等, 2021. 2019年渤海氮磷营养盐季节变化及富营养化状况. 海洋环境科学, 40(6): 915-921 |
匡翠萍, 俞露露, 顾杰, 等, 2019. 人工岛对金梦海湾水体交换的影响. 中国环境科学, 39(2): 757-767 DOI:10.3969/j.issn.1000-6923.2019.02.039 |
刘东艳, 孙军, 陈宗涛, 等, 2002. 不同氮磷比对中肋骨条藻生长特性的影响. 海洋湖沼通报, (2): 39-44 DOI:10.3969/j.issn.1003-6482.2002.02.006 |
刘述锡, 樊景凤, 王真良, 2013. 北黄海浮游植物群落季节变化. 生态环境学报, 22(7): 1173-1181 DOI:10.3969/j.issn.1674-5906.2013.07.014 |
刘素娟, 陶建华, 赵海萍, 2007. 渤海湾浮游植物的多样性分析. 河北工程大学学报(自然科学版), 24(1): 74-77 DOI:10.3969/j.issn.1673-9469.2007.01.020 |
许歆, 2017. 秦皇岛近海浮游植物群落结构变化及其组学研究[D]. 青岛: 中国科学院大学(中国科学院海洋研究所): 1-144.
|
孙军, 刘东艳, 2005. 2000年秋季渤海的网采浮游植物群落. 海洋学报, 27(3): 124-132 DOI:10.3321/j.issn:0253-4193.2005.03.018 |
孙萍, 李瑞香, 李艳, 等, 2008. 2005年夏末渤海网采浮游植物群落结构. 海洋科学进展, 26(3): 354-363 DOI:10.3969/j.issn.1671-6647.2008.03.011 |
吴玉霖, 孙松, 张永山, 等, 2004. 胶州湾浮游植物数量长期动态变化的研究. 海洋与湖沼, 35(6): 518-523 DOI:10.3321/j.issn:0029-814X.2004.06.006 |
张万磊, 马新, 张永丰, 等, 2020. 2000~2016年秦皇岛海域赤潮特征分析. 海洋湖沼通报, (5): 48-55 |
张建乐, 张秋丰, 2007. 秦皇岛沿岸浮游植物与赤潮生物的初步研究. 河北渔业, (7): 7-12 DOI:10.3969/j.issn.1004-6755.2007.07.005 |
张海波, 王丽莎, 石晓勇, 等, 2019. 渤海中部营养盐季节变化及限制统计分析. 中国环境科学, 39(4): 1579-1588 DOI:10.3969/j.issn.1000-6923.2019.04.028 |
张雪, 王俊, 马武, 等, 2020. 2014年秋季渤海网采浮游植物群落结构. 海洋学报, 42(8): 89-100 |
陈楠生, 黄海龙, 2021. 中国海洋浮游植物和赤潮物种的生物多样性研究进展(一): 渤海. 海洋与湖沼, 52(2): 346-362 |
金德祥, 陈贞奋, 刘师成, 等, 1965. 温度和盐度对三种海洋浮游硅藻生长繁殖的影响. 海洋与湖沼, 7(4): 374-384 |
河北省自然资源厅, 2019-2020. 河北省海洋灾害公报[R]. 石家庄: 河北省自然资源厅.
|
河北省海洋局, 2000~2018. 河北省海洋环境状况公报[R]. 石家庄: 河北省海洋局.
|
赵相伟, 魏玉秋, 孙军, 等, 2020. 春、夏季秦皇岛海域超微型浮游植物. 海洋学报, 42(2): 106-114 |
俞建銮, 李瑞香, 1993. 渤海、黄海浮游植物生态的研究. 黄渤海海洋, 11(3): 52-59 |
栾青杉, 康元德, 王俊, 2018. 渤海浮游植物群落的长期变化(1959~2015). 渔业科学进展, 39(4): 9-18 |
郭术津, 李彦翘, 张翠霞, 等, 2014. 渤海浮游植物群落结构及与环境因子的相关性分析. 海洋通报, 33(1): 95-105 |
曹新宇, 李远眉, 王伟, 等, 2021. 温度和盐度对夜光藻种群密度增长的影响. 大连海洋大学学报, 36(1): 95-102 |
谢琳萍, 孙霞, 王保栋, 等, 2012. 渤黄海营养盐结构及其潜在限制作用的时空分布. 海洋科学, 36(9): 45-53 |
慕建东, 郑向荣, 赵振良, 等, 2015. 秦皇岛海域赤潮期间浮游植物的生态特征. 中国水产科学, 22(2): 288-301 |
霍文毅, 俞志明, 邹景忠, 等, 2001. 胶州湾中肋骨条藻赤潮与环境因子的关系. 海洋与湖沼, 32(3): 311-318 |
CHARLON R J, LOVELOCK J E, ANDREAE M O, et al, 1987. Oceanic phytoplankton, atmospheric Sulphur, cloud albedo and climate. Nature, 326(6114): 655-661 |
DONG Y L, ZUO L M, MA W, et al, 2021. Phytoplankton community organization and succession by sea warming: a case study in thermal discharge area of the northern coastal seawater of China. Marine Pollution Bulletin, 169: 112538 |
DUFRENE M, LEGENDRE P, 1997. Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecological Monographs, 67(3): 345-366 |
EGGE J K, 1998. Are diatoms poor competitors at low phosphate concentrations?. Journal of Marine Systems, 16(3/4): 191-198 |
FIELD J G, CLARKE K R, WARWICK R M, 1982. A practical strategy for analysing multispecies distribution patterns. Marine Ecology Progress Series, 8(1): 37-52 |
PADISÁK J, BORICS G, GRIGORSZKY I, et al, 2006. Use of phytoplankton assemblages for monitoring ecological status of lakes within the water framework directive: the assemblage index. Hydrobiologia, 553(1): 1-14 |
PIELOU E C, 1969. An Introduction to Mathematical Ecology. New York, USA: Wiley, 286
|
RIEGMAN R, DE BOER M, DE SENERPONT DOMIS L, 1996. Growth of harmful marine algae in multispecies cultures. Journal of Plankton Research, 18(10): 1851-1866 |
SHANNON C E, WEAVER W, 1949. The Mathematical Theory of Communication. Urbana: University of Illinois Press, 117
|
SUIKKANEN S, LAAMANEN M, HUTTUNEN M, 2007. Long-term changes in summer phytoplankton communities of the open northern Baltic Sea. Estuarine, Coastal and Shelf Science, 71(3/4): 580-592 |
YU R C, ZHANG Q C, LIU Y, et al, 2021. The dinoflagellate Alexandrium catenella producing only carbamate toxins may account for the seafood poisonings in Qinhuangdao, China. Harmful Algae, 103: 101980 |