 2023, Vol. 54
2023, Vol. 54中国海洋湖沼学会主办。
文章信息
- 张涛, 平洪领, 史会来, 付铁中, 卢斌, 何杰, 李彬. 2023.
- ZHANG Tao, PING Hong-Ling, SHI Hui-Lai, FU Tie-Zhong, LU Bin, HE Jie, LI Bin. 2023.
- 不同盐度梯度和规格体重对横带髭鲷(Hapalogenys mucronatus)幼鱼耗氧率(RO)及排氨率(RN)的影响
- INFLUENCE OF SALINITY AND BODY WEIGHT ON OXYGEN CONSUMPTION AND AMMONIA EXCRETION OF JUVENILE HAPALOGENYS MUCRONATUS
- 海洋与湖沼, 54(2): 566-572
- Oceanologia et Limnologia Sinica, 54(2): 566-572.
- http://dx.doi.org/10.11693/hyhz20220700186
文章历史
-
收稿日期:2022-07-13
收修改稿日期:2022-08-26




横带髭鲷(Hapalogenys mucronatus)俗称十六枚, 隶属鲈形目(Perciforms)、石鲈科(Pomadasyidae)、髭鲷属(Hapalogenys), 为近海岛礁性鱼类, 具有较高的营养价值、观赏价值和经济价值, 深受江浙沪消费者和垂钓爱好者的青睐。20世纪末, 随着渔业资源的过度捕捞和临港工业的快速发展, 横带髭鲷野生资源量大幅下降, 市场上一度供不应求。横带髭鲷具有广盐、耐低温、恋礁等特性, 不仅是广大养殖户向往的养殖对象, 同时也是渔业资源生态修复的优良品种。长期以来, 由于野生资源匮乏、亲鱼培育成活率低等原因, 横带髭鲷人工繁育一直处于停滞状态, 养殖户主要以收购野生苗种进行零星养殖, 极大制约了产业规模的发展(张涛等, 2020; 平洪领等, 2021)。本团队自2016年开始收集横带髭鲷野生幼鱼, 近年来开展了人工驯养、生殖调控、催产、孵化、苗种培育、病害防治等系列研究, 初步建立了横带髭鲷苗种繁育及养殖技术体系, 但规模化人工繁育技术尚未完全突破。
呼吸与排泄是动物新陈代谢活动的重要生理指标, 也是研究动物能量学、养殖容量和评价动物对其所处生态系统影响的重要内容之一(仇登高等, 2017; 张雨等, 2021)。其中, 耗氧率和排氨率是动物呼吸和排泄代谢的两个重要指标, 可直接或者间接地反映其生长代谢规律、特点及生理生存状况等(黄洋等, 2013; 刘旭佳等, 2017)。鱼类的耗氧率和排氨率除了受自身的状况(体重、发育期等)影响外, 还受外部环境因子(盐度、温度、光照、pH、密度及饵料等)的影响(李加儿等, 2014; 崔前进等, 2018)。目前国内外对于横带髭鲷的研究相对较少, 仅见于分类地位(徐田军等, 2010; Hata et al, 2016)、遗传学分析(喻子牛等, 1994; 王世锋等, 2008; Zheng et al, 2020)、亲体培育(许文军等, 2002; Soh et al, 2005; 姚海富等, 2006)、早期生长发育(史海东等, 2004; 平洪领等, 2021; 付铁中等, 2022)及营养学分析(张涛等, 2020)等研究, 而对横带髭鲷幼鱼呼吸代谢方面的研究尚未见报道。本研究采用室内实验生态学方法, 对横带髭鲷幼鱼在不同盐度梯度及规格体重下的耗氧率(RO)和排氨率(RN)进行了测定分析, 旨在探讨盐度和体重对其呼吸和排泄的生理反应机制, 为横带髭鲷生理生态学研究提供基础数据, 以期对横带髭鲷规模化人工繁育技术的突破、养殖生产及饲料营养配置等提供理论指导。
1 材料与方法 1.1 实验材料实验在浙江省海洋水产研究所西闪试验场完成, 实验用横带髭鲷为本试验场自繁子一代苗种。选择无病无伤、行动迅速、生命力强的个体作为试验用鱼, 按照体重大小分为小(S)、中(M)、大(L)三组(见表 1, 图 1), 分别挑选90尾暂养在3个7 m3水泥池中, 暂养时间为3 d。实验用水为经暗沉淀和砂滤后的自然海水, 每天吸污换水各1次, 充气微流水养殖; 饵料为舟山近海拖网葛氏长臂虾, 将虾肉剪碎投喂, 每天投喂两次(08:00, 17:00); 暂养期间水温为(26.5±0.5) ℃, pH为8.2±0.2, 盐度为23.2±0.4。
| 试验组 | 体重/g | 全长/cm |
| S | 1.55±0.37 | 4.46±0.36 |
| M | 3.92±0.74 | 5.91±0.31 |
| L | 9.08±1.38 | 7.62±0.37 |
 |
| 图 1 不同规格横带髭鲷实验幼鱼 Fig. 1 Different body sizes of juvenile H. mucronatus used for experiment |
实验设6个盐度梯度, 分别为10、15、20、25、30和35, 采用砂滤海水与自来水和粗海盐的方式调配而成。将3种规格实验用鱼随机选取15尾分别暂养至6个200 L聚丙烯(pp)塑料桶中, 每天降低或提高盐度2~3, 达到实验设定盐度后再暂养7 d开始实验, 暂养期间养殖管理同1.1, 实验前停食24 h。
1.3 耗氧率和排氨率的测定实验装置为自制静水密封式呼吸测定仪, 根据鱼体大小选用6.5 L塑料桶作为实验容器, 塑料桶用塑料薄膜封口, 保证无气泡。每个盐度每种规格均设3个塑料桶作为实验组, 每桶中放3尾鱼, 每个盐度组另设1个无鱼的塑料桶作为空白对照组。塑料桶密闭1.5 h后, 用溶解氧测量仪现场测定实验桶和空白桶中水体溶解氧。溶解氧测定后, 利用虹吸法在每个实验桶各取200 mL水样于实验室中利用HACH分光光度计(DR/2010)测定水中氨氮含量。每组实验结束后均精确测定该组实验鱼的体重、全长等生物学数据。
1.4 相关系数及计算方法根据实验组和对照组水体中溶解氧、氨氮含量的变化, 按下列公式计算横带髭鲷幼鱼耗氧率(RO)、排氨率(RN)以及氧氮比(O: N):
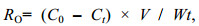 (1)
(1)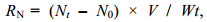 (2)
(2)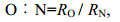 (3)
(3)式中: RO为耗氧率[单位: mg/(g·h)]; RN为排氨率[单位: mg/(g·h)]; C0、Ct分别为实验结束时对照组和实验组中的溶解氧浓度(单位: mg/L); N0、Nt分别为实验结束时对照组和实验组中的氨氮质量浓度(单位: mg/L); V为塑料桶体积(单位: L); W为实验鱼的体重(单位: g); t为实验持续时间(单位: h)。
1.5 数据统计与分析所有实验数据均采用Excel软件进行整理统计, 并采用SPSS 20.0软件进行单因素方差分析, 以Duncan多重比较方法检验组间差异, 以P < 0.05为差异显著。
2 结果与分析 2.1 不同盐度梯度及规格体重对横带髭鲷幼鱼耗氧率(RO)的影响如图 2所示, 盐度对三种规格横带髭鲷幼鱼的耗氧率有较为显著的影响, 同一规格下均随着盐度的升高呈先上升后下降的趋势, 均在盐度20时耗氧率达到相对最大值, 均显著高于盐度10时耗氧率(P < 0.05); 其余盐度间耗氧率差异不显著(P > 0.05)。在同一盐度下, 随着横带髭鲷幼鱼体重的增加, 其单位体重的耗氧率逐渐降低; 三种规格横带髭鲷幼鱼的耗氧率均表现为S组显著高于M组和L组(P < 0.05), M组和L组差异不显著(P > 0.05); 耗氧率的变化范围在0.209~0.550 mg/(g·h)之间, 平均为0.332 mg/(g·h)。
 |
| 图 2 不同盐度梯度及规格体重对横带髭鲷幼鱼耗氧率(RO)的影响 Fig. 2 Influences of different salinity and weight sizes on RO of juvenile H. mucronatus |
对横带髭鲷幼鱼的耗氧率(RO)与体重(W)的回归关系进行拟合, 最符合一元三次函数关系: y=a+bx+ cx2+dx3 (a、b、c和d均为常数, x为体重, y为耗氧率), 详见表 2。表明横带髭鲷幼鱼体重与耗氧率呈负相关关系, 随着体重的增加, 耗氧率呈下降的趋势。
| 盐度 | a | b | c | d | F | P | R2 |
| 10 | 0.461 | –0.016 | 0.000 | 1.824×10–5 | 3.981 | 0.086 | 0.705 |
| 15 | 0.691 | –0.050 | 0.002 | –1.821×10–5 | 14.444 | 0.007 | 0.897 |
| 20 | 0.799 | –0.065 | 0.003 | –4.648×10–5 | 11.227 | 0.012 | 0.871 |
| 25 | 0.693 | –0.050 | 0.002 | –1.447×10–5 | 17.090 | 0.005 | 0.911 |
| 30 | 0.700 | –0.058 | 0.002 | –2.614×10–5 | 20.149 | 0.003 | 0.924 |
| 35 | 0.513 | –0.036 | 0.001 | –1.828×10–5 | 10.803 | 0.013 | 0.866 |
方差分析(ANOVA)显示(表 3), 盐度和体重对耗氧率有显著影响(P < 0.05), 而盐度和体重的交互作用对耗氧率影响不显著(P > 0.05)。
| 变异来源 | 自由度 (K) |
平方和 (SSR) |
均方差 (MSR) |
检验量 (F) |
显著性概率 (P) |
| 盐度 | 5 | 0.068 | 0.014 | 6.386 | 0.000 |
| 体重 | 2 | 0.495 | 0.248 | 117.114 | 0.000 |
| 盐度×体重 | 10 | 0.014 | 0.001 | 0.650 | 0.761 |
| 误差 | 36 | 0.076 | 0.002 | ||
| 总变异 | 54 | 6.620 |
如图 3所示, 盐度对三种规格横带髭鲷幼鱼的排氨率有较为显著的影响, 同一规格下均随着盐度的升高呈先上升后下降的趋势, 其排氨率均在盐度25时达到相对最大值; S组和L组在盐度25和20时排氨率差异不显著(P > 0.05), 均显著高于其余盐度组(P < 0.05); M组各盐度间排氨率差异不显著(P > 0.05)。在同一盐度下, 随着横带髭鲷幼鱼体重的增加, 其单位体重的排氨率逐渐降低; 除盐度35外, 三种规格横带髭鲷幼鱼排氨率均表现为S组和M组、M组和L组差异不显著(P > 0.05); 盐度35时, S组排氨率显著高于M组和L组(P < 0.05); 排氨率的变化范围在0.039~0.080 mg/(g·h)之间, 平均为0.055 mg/(g·h)。
 |
| 图 3 不同盐度梯度及规格体重对横带髭鲷幼鱼排氨率(RN)的影响 Fig. 3 Influences of different salinity and body weights on RN of juvenile H. mucronatus |
对横带髭鲷幼鱼的排氨率(RN)与体重(W)的回归关系进行拟合, 最符合幂函数关系: y= exf (e和f均为常数, x为体重, y为排氨率), 详见表 4。表明横带髭鲷幼鱼体重与排氨率呈负相关关系, 随着体重的增加排氨率呈下降的趋势。
| 盐度 | e | f | F | P | R2 |
| 10 | 0.078 | –0.204 | 42.026 | 0.000 | 0.857 |
| 15 | 0.100 | –0.243 | 25.089 | 0.002 | 0.782 |
| 20 | 0.124 | –0.283 | 66.710 | 0.000 | 0.905 |
| 25 | 0.122 | –0.284 | 59.665 | 0.000 | 0.895 |
| 30 | 0.078 | –0.168 | 9.005 | 0.020 | 0.563 |
| 35 | 0.082 | –0.226 | 16.832 | 0.005 | 0.706 |
方差分析(ANOVA)显示(表 5), 盐度和体重对排氨率有显著影响(P < 0.05), 而盐度和体重的交互作用对排氨率影响不显著(P > 0.05)。
| 变异来源 | 自由度 (K) |
平方和 (SSR) |
均方差 (MSR) |
检验量 (F) |
显著性概率 (P) |
| 盐度 | 5 | 0.002 | 0.000 | 11.085 | 0.000 |
| 体重 | 2 | 0.005 | 0.002 | 71.428 | 0.000 |
| 盐度×体重 | 10 | 0.000 | 3.879×10–5 | 1.118 | 0.376 |
| 误差 | 36 | 0.001 | 3.468×10–5 | ||
| 总变异 | 54 | 0.174 |
O: N值为呼吸氧原子数与排出的氨态氮原子数之比, 如表 6、图 4所示, 三种规格横带髭鲷幼鱼O: N值均随盐度的上升呈波动趋势, O: N值波动范围在5.035~7.533之间, 其总的平均值为5.916; 同一规格不同盐度间O: N值差异不显著(P > 0.05), 而三种规格O: N值平均值表现为S组(6.908)显著高于M组(5.426)和L组(5.414) (P < 0.05), L组和M组差异不显著(P > 0.05)。
| 盐度 | S | M | L | ||||||||
| O | N | O: N | O | N | O: N | O | N | O: N | |||
| 10 | 0.395 | 0.056 | 7.099 | 0.251 | 0.050 | 5.035 | 0.209 | 0.039 | 5.337 | ||
| 15 | 0.472 | 0.066 | 7.191 | 0.301 | 0.057 | 5.277 | 0.245 | 0.044 | 5.612 | ||
| 20 | 0.550 | 0.078 | 7.007 | 0.350 | 0.061 | 5.724 | 0.269 | 0.048 | 5.626 | ||
| 25 | 0.498 | 0.080 | 6.203 | 0.321 | 0.062 | 5.183 | 0.248 | 0.049 | 5.080 | ||
| 30 | 0.473 | 0.063 | 7.533 | 0.281 | 0.049 | 5.671 | 0.235 | 0.045 | 5.202 | ||
| 35 | 0.393 | 0.061 | 6.416 | 0.273 | 0.048 | 5.664 | 0.219 | 0.039 | 5.629 | ||
| 平均 | 6.908 | 5.426 | 5.414 | ||||||||
 |
| 图 4 不同盐度下各规格横带髭鲷幼鱼耗氧率(Ro)与排氨率(RN)的比值 Fig. 4 Various ratios of RO to RN of juvenile H. mucronatus at different salinities and weight sizes |
盐度是海水养殖中重要的环境因子之一, 对鱼类的新陈代谢有着重要的影响。呼吸和排泄是鱼类最基础而又最重要的新陈代谢活动之一, 耗氧率和排氨率则是指示鱼类代谢活动变化的最重要指标。有学者提出, 鱼类的标准代谢可分为两个部分: 一是组织的修复与更新所消耗的能量; 二是维持内稳态所消耗的能量(Jobbing, 1985)。
目前, 关于盐度对鱼类呼吸和排泄的影响主要存在两种观点, 一种观点是针对狭盐性鱼类(其适盐范围小), 当处于适盐范围时, 鱼体自身渗透压与外界水环境渗透压相同即达到等渗点时其耗氧率和排氨率均最低, 高于或低于适盐范围时, 其耗氧率和排氨率均升高, 如平鲷(Woo et al, 1995)、珍珠龙胆石斑鱼(刘龙龙等, 2020)、长鳍篮子鱼(章龙珍等, 2009)、钝吻黄盖鲽(崔前进等, 2018)、大泷六线鱼(胡发文等, 2021)等研究结果与此观点相一致, 出现这种现象的主要原因是参照渗透压调节原理, 鱼体在等渗点时渗透压力最小, 此时维持机体内渗透压平衡耗能最少, 代谢率最低, 而远离等渗点时则需要更多的耗能来调节机体渗透压, 代谢率上升(Woo et al, 1995; 闫茂仓等, 2008)。另一种观点是针对广盐性鱼类(其适盐范围大), 当处于适盐范围时其耗氧率和排氨率均最高, 即在一定的盐度范围内鱼体耗氧率和排氨率均随盐度的增加而升高, 达到峰值后, 随盐度的继续增加而降低, 且不同鱼类之间因适盐范围不同而差别较大, 如鮸鱼(闫茂仓等, 2007)、浅色黄姑鱼(李加儿等, 2008)、日本黄姑鱼(柴学军等, 2009)、大黄鱼(郭念岗等, 2014; 张涛等, 2015)、褐菖鮋(严银龙等, 2019)等研究结果与此观点相一致, 出现这种现象的主要原因一部分是由于水体盐度高于或低于鱼体适盐范围时, 鱼体自身的渗透压调节机制受到抑制, 进而表现为鱼体耗氧率和排氨率受到抑制; 另外, 渗透压调节耗能只占代谢能量中很小的一部分, 而用于生长活动的代谢耗能则占主导地位, 进而表现为鱼体在适盐范围内耗氧率和排氨率较高(闫茂仓等, 2008; 吴庆元等, 2014)。
本研究结果表明, 盐度可显著影响横带髭鲷幼鱼呼吸和排泄等新陈代谢活动(P < 0.05), 耗氧率和排氨率均随盐度的增加而升高, 并在盐度20时耗氧率达到峰值、盐度25时排氨率达到峰值, 后均随着盐度的继续增加而降低, 这一结果与上述观点二相一致, 只是不同鱼类间耗氧率和排氨率达到峰值的对应盐度不同, 作者认为这可能与试验动物的物种差异、规格大小及其他环境因子等有关系。对于横带髭鲷而言, 其适盐范围较广, 在适盐范围内适当降低盐度来降低代谢率, 可以达到提高生长速度的目的; 利用其耐低盐的特性, 短时间内在低盐海水(或淡水)中浸浴, 可以达到杀灭寄生虫并防止寄生虫加剧感染的目的。
3.2 不同规格体重对横带髭鲷幼鱼呼吸和排泄的影响规格体重也是影响鱼类呼吸和排泄的一个重要影响因子。大量研究表明, 体重对鱼类耗氧率和排氨率具有显著差异影响(P < 0.05), 如尼罗罗非鱼(Yamamoto, 1992)、金枪鱼(Herrmann et al, 2000)、浅色黄姑鱼(李加儿等, 2008)、条石鲷(闫茂仓等, 2008)、大弹涂鱼(周银环等, 2017)、钝吻黄盖鲽(崔前进等, 2018)、禾花鲤(王丽群等, 2020)等, 这些研究结果均表明鱼体耗氧率和排氨率与体重之间呈负相关关系, 均随体重的增加呈逐渐降低的趋势。
本研究结果表明, 在盐度恒定时, 规格对横带髭鲷幼鱼耗氧率和排氨率影响显著(P < 0.05), 均随着体重的增加而逐渐降低, 研究结果与上述观点一致。相关研究表明(姜祖辉等, 1999; 周银环等, 2017; 刘建勇等, 2019), 水生动物用来维持其生命活动的组织和器官(如肝胰脏和肾脏等)的新陈代谢活力要高于其他非直接维持生命活动的组织和器官(如脂肪和肌肉等); 在水生动物个体较小时, 肝胰脏和肾脏所占总体重的比例较大, 其耗氧率和排氨率较高, 但随着水生动物个体的进一步增大, 肌肉和脂肪等所占总体重的比例要远大于肝胰脏和肾脏等所占比例, 虽然耗氧量和排氨量随着总体重的增加而增加, 但耗氧率和排氨率却随总体重的增加呈现逐渐降低的趋势。规格体重对耗氧率和排氨率的影响也会因生物种类及其生长发育阶段的不同而不同(王刚等, 2010; 吴文广等, 2013; 王鹏帅等, 2017)。
另外, 本研究结果表明, 在盐度10~35之间, 横带髭鲷幼鱼体重(W)与单位体重的耗氧率之间呈一元三次函数关系, 体重(W)与单位体重的排氨率之间呈幂函数关系。而钝吻黄盖鲽(崔前进等, 2018)在盐度8~32时, 体重(W)与单位体重的耗氧率和排氨率均符合一元一次函数关系; 九孔鲍(刘建勇等, 2019)在盐度21~40时, 管角螺(罗杰等, 2008)在盐度12~32时, 尖紫蛤(黄洋等, 2013)在盐度2~18时, 软体部干质量与单位体重的耗氧率和排氨率之间均符合幂函数关系; 上述研究报道与本研究结果存在一定差异, 作者认为这可能与试验品种、规格大小、盐度范围及营养特征的不同有关。
3.3 氧氮比(O: N)对横带髭鲷幼鱼能源物质的分析O: N值表示生物体内蛋白质、脂肪和碳水化合物分解代谢时所消耗的氧气和排出的氨态氮的比例, 可以作为生物适应环境压力的指标之一, 已被大量研究者用来估计不同生物代谢过程中能源物质的化学本质以及能量消耗情况(焦海峰等, 2015; 刘旭佳等, 2017; 崔前进等, 2018; 刘鹏远等, 2020; 周建聪等, 2022)。有学者认为, 如果机体耗能完全由蛋白质为氧化基质提供, O: N值约为7 (Mayzaud, 1976); 如果机体耗能完全由蛋白质和脂肪氧化供给, O: N值约为24 (Ikeda, 1974); 如果机体耗能主要由脂肪或碳水化合物提供、甚至完全由脂肪或碳水化合物提供能量的话, O: N值将大于24, 甚至无穷大(Conover et al, 1968)。
本研究中, 三种规格横带髭鲷幼鱼O: N值均在适宜盐度25时出现了相对低值, 表明在此盐度时需要消耗较少的能量即可维持体内环境的稳定; 小规格O: N值相对离散, 且显著大于大规格和中规格的O: N值(P < 0.05), 表明小规格需要消耗较大的能量来适应外界水环境盐度的改变; 三种规格O: N的比值范围在5.035~7.533, 说明该实验条件下三种规格横带髭鲷幼鱼的能源物质比例基本一致, 主要以蛋白质作为能源物质。因此, 在横带髭鲷养殖生产过程中, 为保证其较快的生长速度, 需要灵活调整不同生长阶段时饵料的蛋白质、脂肪和碳水化合物的含量。
4 结论综上所述, 横带髭鲷体重与单位体重的耗氧率之间的关系呈一元三次函数关系y=a+bx+cx2+dx3, 体重与单位体重的排氨率之间的关系呈幂函数关系: y=exf, 均呈负相关关系。盐度和体重对横带髭鲷的耗氧率、排氨率均有显著影响(P < 0.05), 但盐度和体重的交互作用对耗氧率、排氨率影响均不显著(P > 0.05)。三种规格横带髭鲷在盐度10~35时主要以蛋白质为能源。
王世锋, 杜佳莹, 苏永全, 等, 2008. 厦门近海横带髭鲷野生群体遗传结构分析. 厦门大学学报(自然科学版), 47(1): 88-93 DOI:10.3321/j.issn:0438-0479.2008.01.020 |
王刚, 李加儿, 区又君, 等, 2010. 卵形鲳鲹幼鱼耗氧率和排氨率的初步研究. 动物学杂志, 45(3): 116-121 DOI:10.13859/j.cjz.2010.03.029 |
王丽群, 张旭, 宿志健, 等, 2020. 体质量和温度对禾花鲤耗氧率与窒息点的影响. 安徽农业科学, 48(10): 87-90 DOI:10.3969/j.issn.0517-6611.2020.10.024 |
王鹏帅, 蒋霞敏, 韩庆喜, 等, 2017. 盐度和温度对不同规格虎斑乌贼幼体的耗氧率、排氨率和窒息点的影响. 水生生物学报, 41(5): 1027-1035 |
仇登高, 黄种持, 郑乐云, 等, 2017. 温度对不同规格珍珠龙胆石斑鱼(棕点石斑鱼♀×鞍带石斑鱼♂)呼吸和排泄的影响. 中国水产科学, 24(2): 295-305 |
平洪领, 张涛, 史会来, 等, 2021. 横带髭鲷早期生长发育特征. 中国水产科学, 28(3): 276-287 |
史海东, 毛国民, 王海岳, 2004. 温度和盐度对横带髭鲷胚胎发育的影响. 上海水产大学学报, 13(3): 230-234 |
付铁中, 平洪领, 张涛, 等, 2022. 横带髭鲷(Hapalogenys mucronatus)仔鱼饥饿不可逆点及摄食生长特性研究. 海洋与湖沼, 53(6): 1494-1502 |
刘龙龙, 罗鸣, 陈傅晓, 等, 2020. 盐度对珍珠龙胆石斑鱼幼鱼渗透调节与耗氧率的影响. 中国水产科学, 27(6): 692-700 |
刘旭佳, 沈夏霜, 黄国强, 等, 2017. 盐度和规格对光裸方格星虫耗氧率及排氨率的影响. 水产学报, 41(1): 64-69 |
刘建勇, 陈园媛, 曹伏君, 等, 2019. 盐度对不同规格九孔鲍耗氧量和排氨率的影响. 广东海洋大学学报, 39(5): 129-134 |
刘鹏远, 张春霞, 赵本良, 等, 2020. 盐胁迫对福寿螺耗氧率和排氨率的影响. 中国生态农业学报(中英文), 28(7): 1072-1078 |
闫茂仓, 单乐州, 邵鑫斌, 等, 2007. 盐度和pH值对鮸鱼幼鱼耗氧率和氨氮排泄率的影响. 台湾海峡, 26(1): 85-91 DOI:10.3969/j.issn.1000-8160.2007.01.011 |
闫茂仓, 单乐州, 谢起浪, 等, 2008. 温度、盐度及体重对条石鲷幼鱼耗氧率和排氨率的影响. 海洋科学进展, 26(4): 486-496 DOI:10.3969/j.issn.1671-6647.2008.04.010 |
许文军, 辛剑, 张学舒, 等, 2002. 横带髭鲷Hapalogeny mucronatus (Eydoux et Souleyet)白点虫病的防治研究. 现代渔业信息, 17(11): 24-25, 13 |
严银龙, 袁新程, 施永海, 等, 2019. 盐度对褐菖鲉幼鱼生长、耗氧率和排氨率的影响. 大连海洋大学学报, 34(4): 545-551 |
李加儿, 刘士瑞, 区又君, 等, 2008. 浅色黄姑鱼幼鱼耗氧率、排氨率及窒息点的初步研究. 海洋学报, 30(5): 165-170 |
李加儿, 曹守花, 区又君, 等, 2014. 温度、盐度和pH对鲻幼鱼耗氧率、排氨率以及窒息点的影响. 中国水产科学, 21(5): 954-962 |
吴文广, 张继红, 方建光, 等, 2013. 盐度对不同规格泥螺耗氧率和排氨率的影响. 生态学杂志, 32(9): 2457-2461 |
吴庆元, 蒋玫, 李磊, 等, 2014. 低盐度胁迫对鲻鱼(Mugil cephalus)幼鱼耗氧率和排氨率的影响. 海洋环境科学, 33(2): 177-181 |
张雨, 陈爱华, 吴杨平, 等, 2021. 大竹蛏Solen grandis耗氧率及排氨率响应面法分析. 水产学杂志, 34(2): 65-70 |
张涛, 王庚申, 平洪领, 等, 2015. 盐度对大黄鱼幼鱼耗氧率和排氨率的影响研究. 浙江海洋学院学报(自然科学版), 34(4): 310-313 |
张涛, 平洪领, 史会来, 等, 2020. 舟山海域不同规格横带髭鲷营养成分分析及品质评价. 浙江海洋大学学报(自然科学版), 39(6): 502-508 |
罗杰, 刘楚吾, 李锋, 等, 2008. 盐度及规格对管角螺耗氧率和排氨率的影响. 海洋科学, 32(5): 46-50 |
周建聪, 蔡利, 杨静茹, 等, 2022. 温度和氨氮对不同规格红螯螯虾耗氧率与排氨率的影响. 渔业科学进展, 43(3): 95-102 |
周银环, 曹伏君, 罗杰, 等, 2017. 不同温度梯度和规格体重对大弹涂鱼(Boleophthalmus pectinirostris)耗氧率(RO)和排氨率(RN)的影响. 海洋与湖沼, 48(2): 392-397 |
胡发文, 王晓龙, 高凤祥, 等, 2021. 温度、盐度和两种麻醉剂对大泷六线鱼幼鱼耗氧率、排氨率的影响. 海洋科学, 45(1): 54-61 |
姜祖辉, 王俊, 唐启升, 1999. 菲律宾蛤仔呼吸和排泄规律的研究. 海洋水产研究, 20(1): 40-44 |
姚海富, 毛国民, 史海东, 2006. 横带髭鲷Hapalogenys mucronatus (Eydoux et Souleyet)亲鱼培育、产卵和孵化的初步试验. 现代渔业信息, 21(4): 15-17 |
柴学军, 胡则辉, 徐君卓, 等, 2009. 盐度和pH对日本黄姑鱼幼鱼耗氧率和排氨率的影响. 浙江海洋学院学报(自然科学版), 28(2): 146-150 |
徐田军, 王健鑫, 孙悦娜, 等, 2010. 基于髭鲷属鱼类Cyt b基因全序列探讨髭鲷属在鲈总科的分类地位. 动物分类学报, 35(3): 530-536 |
郭念岗, 吴雄飞, 沈伟良, 2014. 温度和盐度对岱衢族大黄鱼排氨率和耗氧率的影响. 渔业现代化, 41(3): 21-25 |
黄洋, 黄海立, 林国游, 等, 2013. 盐度、pH和规格对尖紫蛤(Soletellina acuta)耗氧率和排氨率的影响. 海洋与湖沼, 44(1): 120-125 |
崔前进, 尚胜男, 蔡忠璐, 等, 2018. 盐度、温度和体质量对钝吻黄盖鲽幼鱼排氨率和耗氧率的影响. 上海海洋大学学报, 27(1): 64-72 |
章龙珍, 杨金海, 刘鉴毅, 等, 2009. 温度、盐度、pH和麻醉剂对长鳍篮子鱼幼鱼耗氧率的影响. 生态学杂志, 28(8): 1494-1498 |
喻子牛, 孔晓瑜, 徐文武, 等, 1994. 斜带髭鲷Hapalogenys nitens (Richardson)和横带髭鲷H. mucronatus (Eydoux et Souleyet)的核型. 青岛海洋大学学报(自然科学版), 24(2): 175-180 |
焦海峰, 严巧娜, 郑丹, 等, 2015. 温度和盐度对埋栖性双壳类泥蚶(Tegillarca granosa)呼吸、排泄的影响. 海洋与湖沼, 46(6): 1333-1338 |
CONOVER R J, CORNER E D S, 1968. Respiration and nitrogen excretion by some marine zooplankton in relation to their life cycles. Journal of the Marine Biological Association of the United Kingdom, 48(1): 49-75 |
HATA M, MOTOMURA H, 2016. First record of Hapalogenys analis (Perciformes: Haemulidae) from Uchinoura Bay in the east coast of Osumi Peninsula, Kagoshima Prefecture, southern Japan. Nature of Kagoshima, 42: 243-248 |
HERRMANN J P, ENDERS E C, 2000. Effect of body size on the standard metabolism of horse mackerel. Journal of Fish Biology, 57(3): 746-760 |
IKEDA T, 1974. Nutritional ecology of marine zooplankton. Memoirs of the Faculty of Fisheries Hokkaido University, 22(1): 1-97 |
JOBBING M, 1985. Growth and metabolism[M] // TYTLER P, CAKOW P. Fish Energetics: Perspectives. London: Croom Helm: 257-281.
|
MAYZAUD P, 1976. Respiration and nitrogen excretion of zooplankton. Ⅳ. The influence of starvation on the metabolism and the biochemical composition of some species. Marine Biology, 37(1): 47-58 |
SOH H Y, KWAK S N, 2005. Feeding habits of belted beard grunt, Hapalogenys mucronatus, in the coastal waters off Sori Island, Yeosu, Korea. Korean Journal of Ichthyology, 17(4): 258-263 |
WOO N Y S, Kelly S P, 1995. Effects of salinity and nutritional status on growth and metabolism of Spams sarba in a closed seawater system. Aquaculture, 135(1/2/3): 229-238 |
YAMAMOTO K I, 1992. Relationship of respiration to body weight in the tilapia Oreochromis Niloticus under resting and normoxic conditions. Comparative Biochemistry and Physiology Part A: Physiology, 103(1): 81-83 |
ZHENG Y Y, FENG L X, LIU H Y, et al, 2020. The complete mitochondrial genome of Hapalogenys analis (Perciformes, Haemulidea) except for control region, obtained by whole genome sequencing. Mitochondrial DNA Part B, 5(3): 2807-2808 |